

Trans-synaptic spread of tau pathology in vivo
- PMID: 22312444
- PMCID: PMC3270029
- DOI: 10.1371/journal.pone.0031302
Trans-synaptic spread of tau pathology in vivo
Abstract
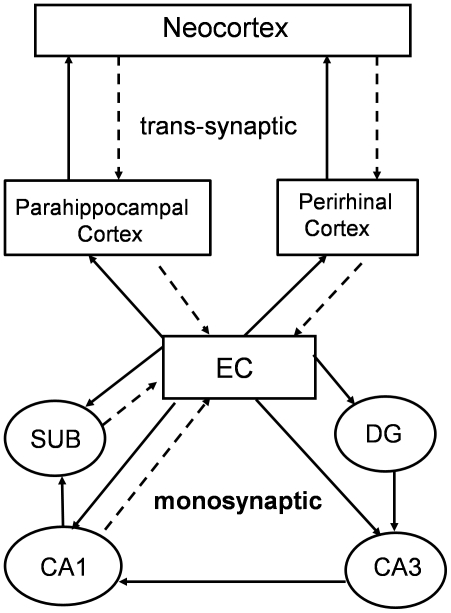
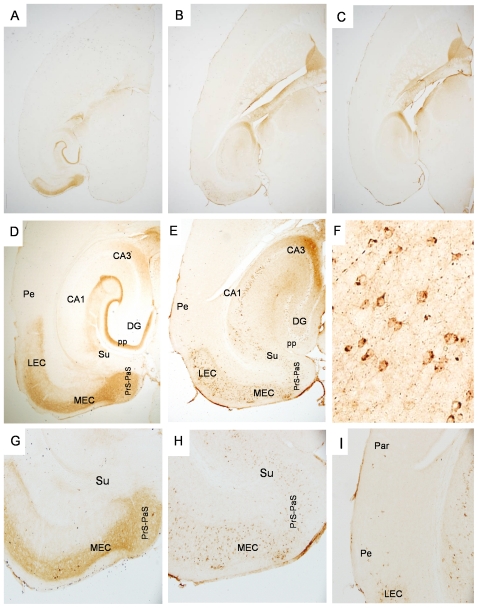
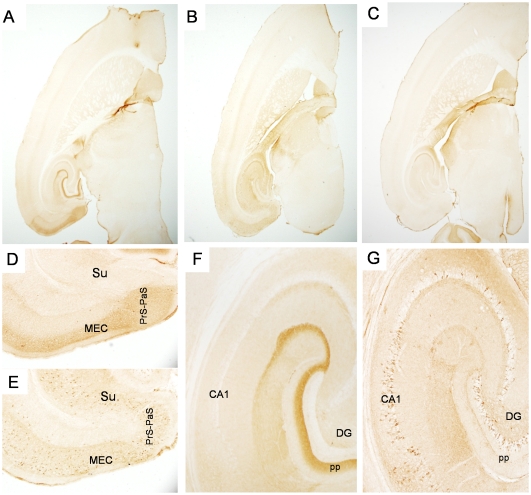
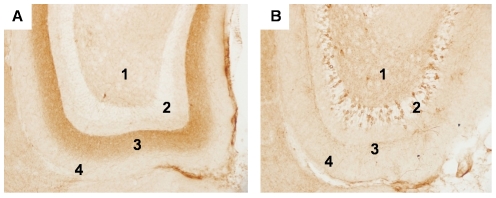
Tauopathy in the brain of patients with Alzheimer's disease starts in the entorhinal cortex (EC) and spreads anatomically in a defined pattern. To test whether pathology initiating in the EC spreads through the brain along synaptically connected circuits, we have generated a transgenic mouse model that differentially expresses pathological human tau in the EC and we have examined the distribution of tau pathology at different timepoints. In relatively young mice (10-11 months old), human tau was present in some cell bodies, but it was mostly observed in axons within the superficial layers of the medial and lateral EC, and at the terminal zones of the perforant pathway. In old mice (>22 months old), intense human tau immunoreactivity was readily detected not only in neurons in the superficial layers of the EC, but also in the subiculum, a substantial number of hippocampal pyramidal neurons especially in CA1, and in dentate gyrus granule cells. Scattered immunoreactive neurons were also seen in the deeper layers of the EC and in perirhinal and secondary somatosensory cortex. Immunoreactivity with the conformation-specific tau antibody MC1 correlated with the accumulation of argyrophilic material seen in old, but not young mice. In old mice, axonal human tau immunoreactivity, especially at the endzones of the perforant pathway, was greatly reduced. Relocalization of tau from axons to somatodendritic compartments and propagation of tauopathy to regions outside of the EC correlated with mature tangle formation in neurons in the EC as revealed by thioflavin-S staining. Our data demonstrate propagation of pathology from the EC and support a trans-synaptic mechanism of spread along anatomically connected networks, between connected and vulnerable neurons. In general, the mouse recapitulates the tauopathy that defines the early stages of AD and provides a model for testing mechanisms and functional outcomes associated with disease progression.
Conflict of interest statement
Figures
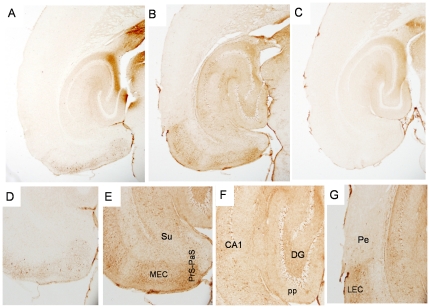
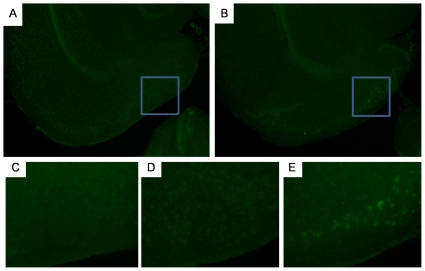
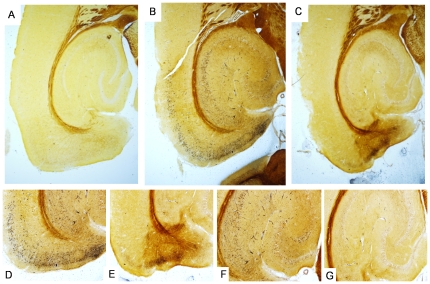
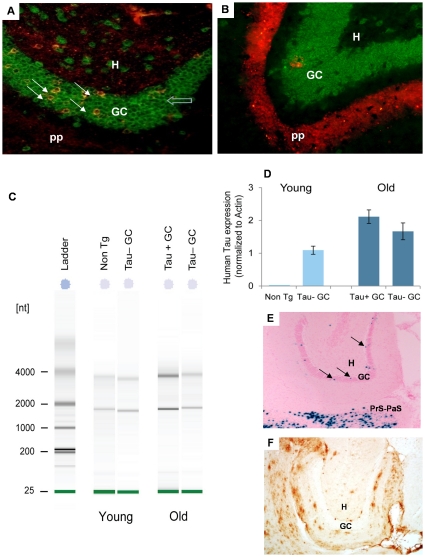
References
-
- Braak H, Braak E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991;82:239–259. - PubMed
-
- van Groen T, Miettinen P, Kadish I. The entorhinal cortex of the mouse: organization of the projection to the hippocampal formation. Hippocampus. 2003;13:133–149. - PubMed
-
- Witter MP, Wouterlood FG, Naber PA, Van Haeften T. Anatomical organization of the parahippocampal-hippocampal network. Ann N Y Acad Sci. 2000;911:1–24. - PubMed
-
- Braak H, Del Tredici K. Alzheimer's pathogenesis: is there neuron-to-neuron propagation? Acta Neuropathol. 2011;121:589–595. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
