

TAK-1/p38/nNFκB signaling inhibits myoblast differentiation by increasing levels of Activin A
- PMID: 22313861
- PMCID: PMC3295657
- DOI: 10.1186/2044-5040-2-3
TAK-1/p38/nNFκB signaling inhibits myoblast differentiation by increasing levels of Activin A
Abstract
Background: Skeletal-muscle differentiation is required for the regeneration of myofibers after injury. The differentiation capacity of satellite cells is impaired in settings of old age, which is at least one factor in the onset of sarcopenia, the age-related loss of skeletal-muscle mass and major cause of frailty. One important cause of impaired regeneration is increased levels of transforming growth factor (TGF)-β accompanied by reduced Notch signaling. Pro-inflammatory cytokines are also upregulated in aging, which led us hypothesize that they might potentially contribute to impaired regeneration in sarcopenia. Thus, in this study, we further analyzed the muscle differentiation-inhibition pathway mediated by pro-inflammatory cytokines in human skeletal muscle cells (HuSKMCs).
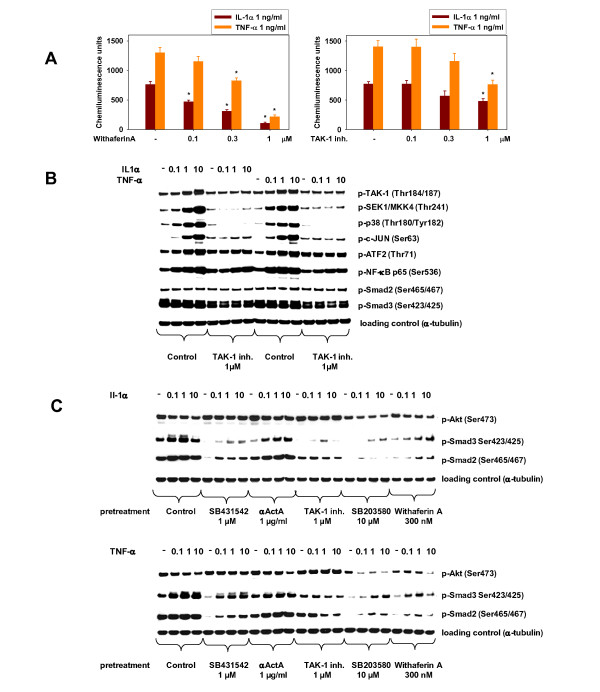
Methods: We studied the modulation of HuSKMC differentiation by the pro-inflammatory cytokines interleukin (IL)-1α and tumor necrosis factor (TNF)-α The grade of differentiation was determined by either imaging (fusion index) or creatine kinase (CK) activity, a marker of muscle differentiation. Secretion of TGF-β proteins during differentiation was assessed by using a TGF-β-responsive reporter-gene assay and further identified by means of pharmacological and genetic inhibitors. In addition, signaling events were monitored by western blotting and reverse transcription PCR, both in HuSKMC cultures and in samples from a rat sarcopenia study.
Results: The pro-inflammatory cytokines IL-1α and TNF-α block differentiation of human myoblasts into myotubes. This anti-differentiation effect requires activation of TGF-β-activated kinase (TAK)-1. Using pharmacological and genetic inhibitors, the TAK-1 pathway could be traced to p38 and NFκB. Surprisingly, the anti-differentiation effect of the cytokines required the transcriptional upregulation of Activin A, which in turn acted through its established signaling pathway: ActRII/ALK/SMAD. Inhibition of Activin A signaling was able to rescue human myoblasts treated with IL-1β or TNF-α, resulting in normal differentiation into myotubes. Studies in aged rats as a model of sarcopenia confirmed that this pro-inflammatory cytokine pathway identified is activated during aging.
Conclusions: In this study, we found an unexpected connection between cytokine and Activin signaling, revealing a new mechanism by which cytokines affect skeletal muscle, and establishing the physiologic relevance of this pathway in the impaired regeneration seen in sarcopenia.
Figures





References
-
- Coleman ME, DeMayo F, Yin KC, Lee HM, Geske R, Montgomery C, Schwartz RJ. Myogenic vector expression of insulin-like growth factor I stimulates muscle cell differentiation and myofiber hypertrophy in transgenic mice. J Biol Chem. 1995;270(20):12109–12116. doi: 10.1074/jbc.270.20.12109. - DOI - PubMed
-
- Florini JR, Ewton DZ, Coolican SA. Growth hormone and the insulin-like growth factor system in myogenesis. Endocr Rev. 1996;17(5):481–517. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
