

SHAPE-directed discovery of potent shRNA inhibitors of HIV-1
- PMID: 22314289
- PMCID: PMC3321596
- DOI: 10.1038/mt.2011.299
SHAPE-directed discovery of potent shRNA inhibitors of HIV-1
Abstract
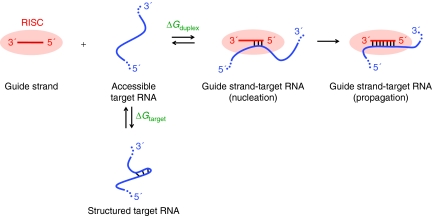
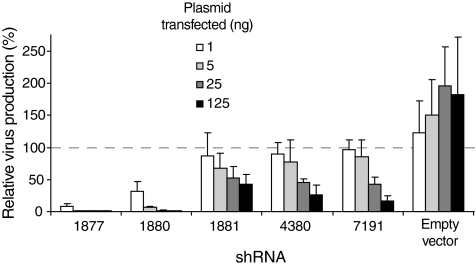
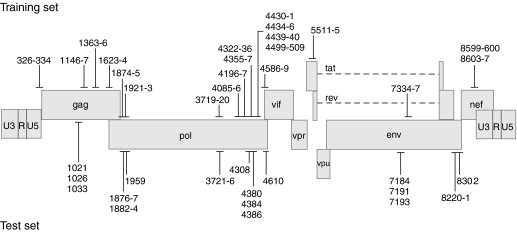
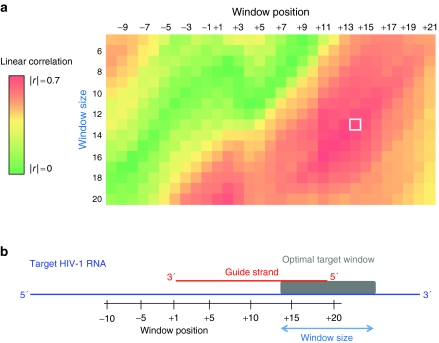
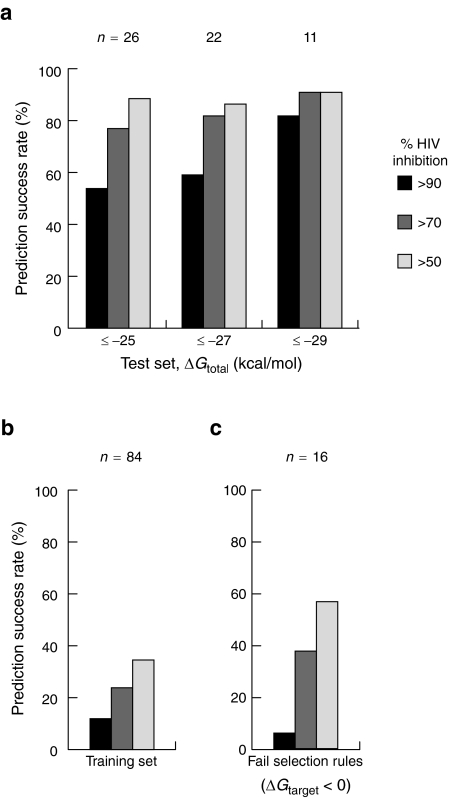
The RNA interference (RNAi) pathway can be exploited using short hairpin RNAs (shRNAs) to durably inactivate pathogenic genes. Prediction of optimal target sites is notoriously inaccurate and current approaches applied to HIV-1 show weak correlations with virus inhibition. In contrast, using a high-content model for disrupting pre-existing intramolecular structure in the HIV-1 RNA, as achievable using high-resolution SHAPE (selective 2'-hydroxyl acylation analyzed by primer extension) chemical probing information, we discovered strong correlations between inhibition of HIV-1 production in a quantitative cell-based assay and very simple thermodynamic features in the target RNA. Strongest inhibition occurs at RNA target sites that both have an accessible "seed region" and, unexpectedly, are structurally accessible in a newly identified downstream flanking sequence. We then used these simple rules to create a new set of shRNAs and achieved inhibition of HIV-1 production of 90% or greater for up to 82% of designed shRNAs. These shRNAs inhibit HIV-1 replication in therapy-relevant T cells and show no or low cytotoxicity. The remarkable success of this straightforward SHAPE-based approach emphasizes that RNAi is governed, in significant part, by very simple, predictable rules reflecting the underlying RNA structure and illustrates principles likely to prove broadly useful in understanding transcriptome-scale biological recognition and therapeutics involving RNA.
Figures

References
-
- Cruz JA., and, Westhof E. The dynamic landscapes of RNA architecture. Cell. 2009;136:604–609. - PubMed
-
- Mello CC., and, Conte D., Jr Revealing the world of RNA interference. Nature. 2004;431:338–342. - PubMed
-
- Wu L., and, Belasco JG. Let me count the ways: mechanisms of gene regulation by miRNAs and siRNAs. Mol Cell. 2008;29:1–7. - PubMed
-
- Wadhwa R, Kaul SC, Miyagishi M., and, Taira K. Vectors for RNA interference. Curr Opin Mol Ther. 2004;6:367–372. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
