

Draft genome of the pearl oyster Pinctada fucata: a platform for understanding bivalve biology
- PMID: 22315334
- PMCID: PMC3325083
- DOI: 10.1093/dnares/dss005
Draft genome of the pearl oyster Pinctada fucata: a platform for understanding bivalve biology
Abstract
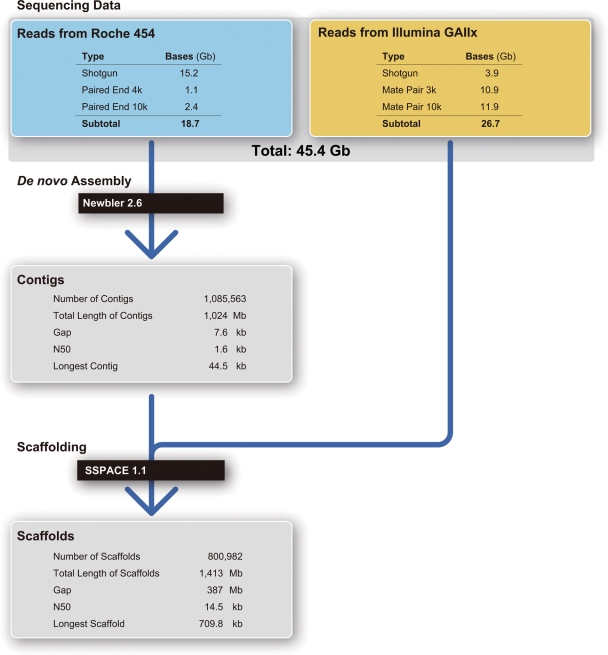
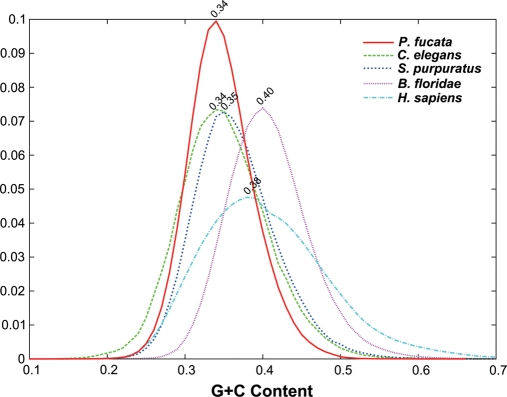
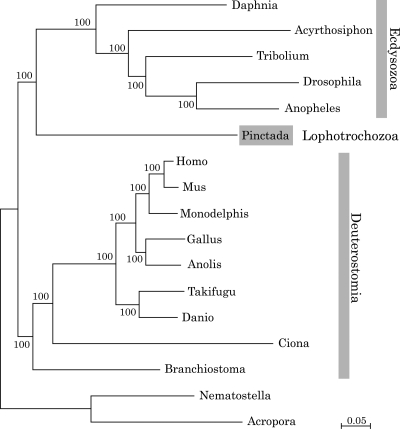
The study of the pearl oyster Pinctada fucata is key to increasing our understanding of the molecular mechanisms involved in pearl biosynthesis and biology of bivalve molluscs. We sequenced ~1150-Mb genome at ~40-fold coverage using the Roche 454 GS-FLX and Illumina GAIIx sequencers. The sequences were assembled into contigs with N50 = 1.6 kb (total contig assembly reached to 1024 Mb) and scaffolds with N50 = 14.5 kb. The pearl oyster genome is AT-rich, with a GC content of 34%. DNA transposons, retrotransposons, and tandem repeat elements occupied 0.4, 1.5, and 7.9% of the genome, respectively (a total of 9.8%). Version 1.0 of the P. fucata draft genome contains 23 257 complete gene models, 70% of which are supported by the corresponding expressed sequence tags. The genes include those reported to have an association with bio-mineralization. Genes encoding transcription factors and signal transduction molecules are present in numbers comparable with genomes of other metazoans. Genome-wide molecular phylogeny suggests that the lophotrochozoan represents a distinct clade from ecdysozoans. Our draft genome of the pearl oyster thus provides a platform for the identification of selection markers and genes for calcification, knowledge of which will be important in the pearl industry.
Figures

References
-
- Weiss I.M., Tuross N., Addadi L., Weiner S. Mollusc larval shell formation: amorphous calcium carbonate is a precursor phase for aragonite. J. Exp. Zool. 2002;293:478–91. doi:10.1002/jez.90004. - DOI - PubMed
-
- Addadi L., Raz S., Weiner S. Taking advantage of disorder: amorphous calcium carbonate and its roles in biomineralization. Adv. Mater. 2003;15:959–70. doi:10.1002/adma.200300381. - DOI
-
- Gerhke N., Nassif N., Pinna N., Antonetti M., Gupta H.S. Retrosynthesis of nacre via amorphous precursor particles. Chem. Mater. 2005;17:6514–6. doi:10.1021/cm052150k. - DOI
-
- Marin F., Luquet G., Marie B., Medakovic D. Molluscan shell proteins: primary structure, origin, and evolution. Curr. Top. Dev. Biol. 2008;80:209–76. doi:10.1016/S0070-2153(07)80006-8. - DOI - PubMed
-
- Sarashina I., Endo K. Skeletal matrix proteins of invertebrate animals: comparative analysis of their amino acid sequences. Paleontol. Res. 2006;10:311–36. doi:10.2517/prpsj.10.311. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
