

RNA synthesis by the brome mosaic virus RNA-dependent RNA polymerase in human cells reveals requirements for de novo initiation and protein-protein interaction
- PMID: 22318148
- PMCID: PMC3318653
- DOI: 10.1128/JVI.00069-12
RNA synthesis by the brome mosaic virus RNA-dependent RNA polymerase in human cells reveals requirements for de novo initiation and protein-protein interaction
Abstract
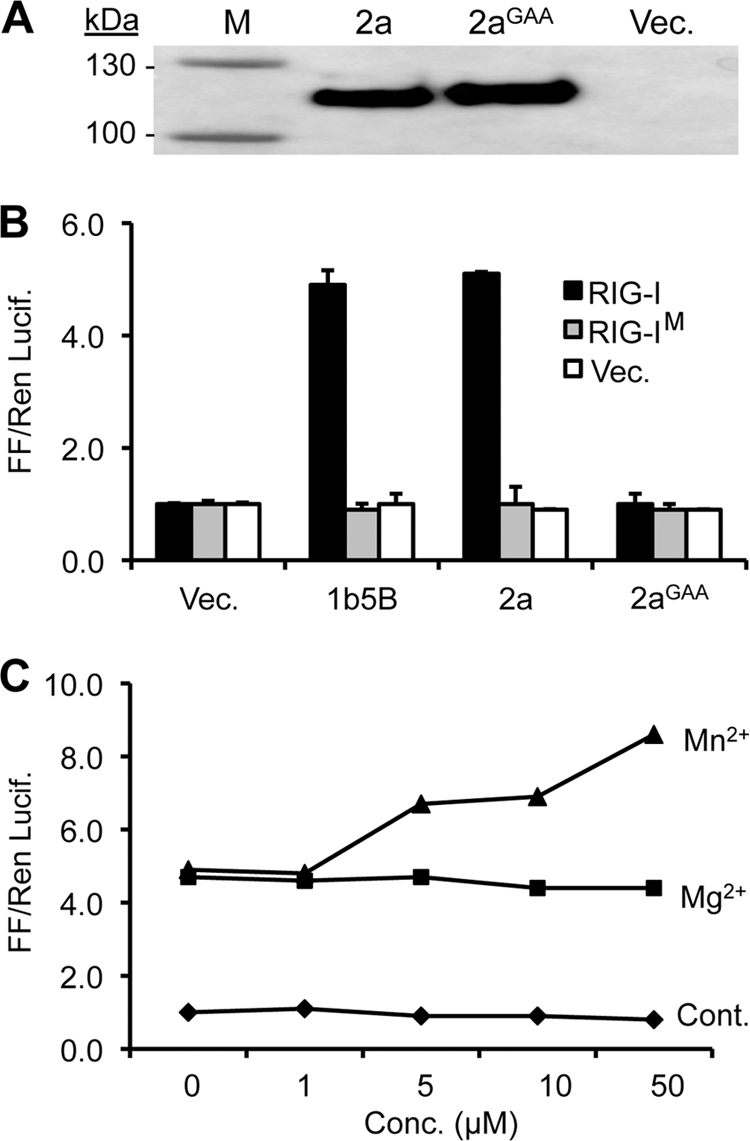
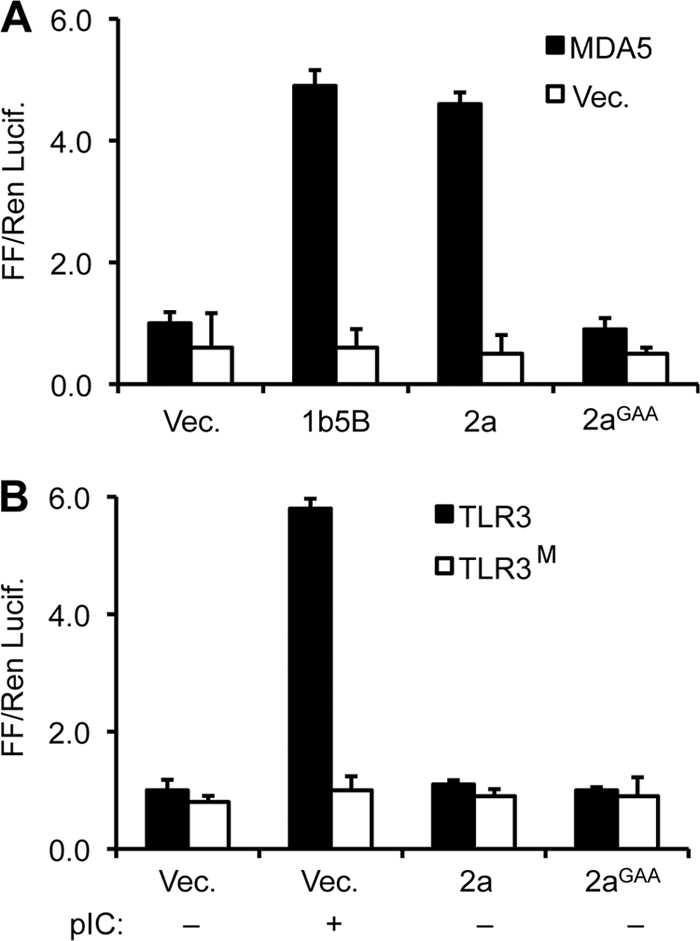
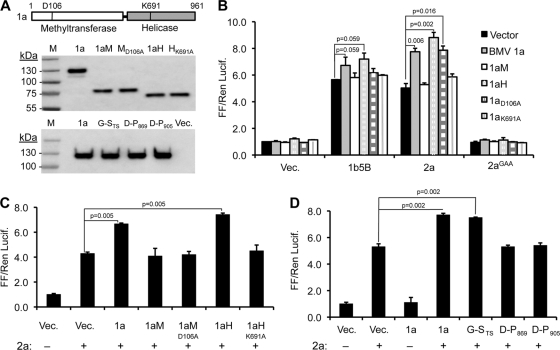
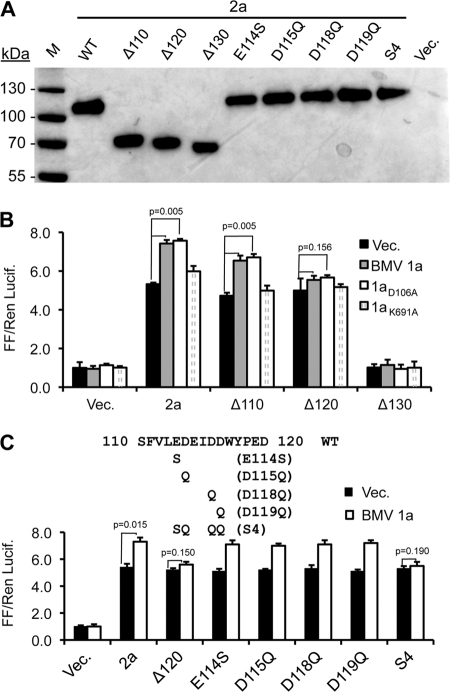
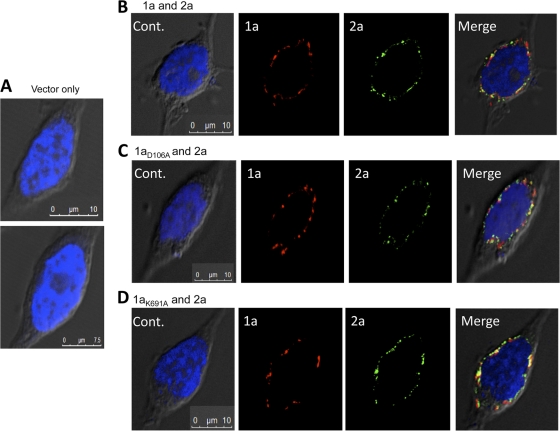
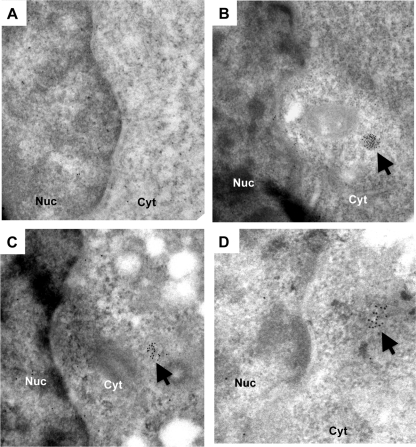
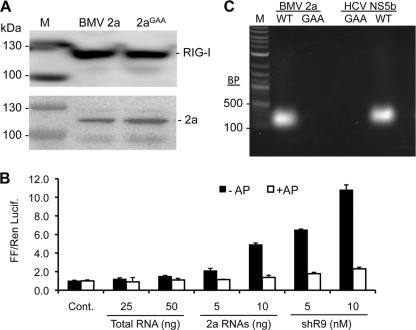
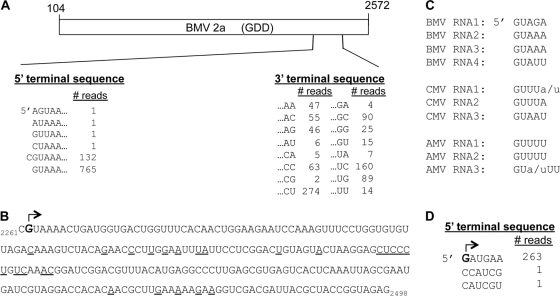
Brome mosaic virus (BMV) is a model positive-strand RNA virus whose replication has been studied in a number of surrogate hosts. In transiently transfected human cells, the BMV polymerase 2a activated signaling by the innate immune receptor RIG-I, which recognizes de novo-initiated non-self-RNAs. Active-site mutations in 2a abolished RIG-I activation, and coexpression of the BMV 1a protein stimulated 2a activity. Mutations previously shown to abolish 1a and 2a interaction prevented the 1a-dependent enhancement of 2a activity. New insights into 1a-2a interaction include the findings that helicase active site of 1a is required to enhance 2a polymerase activity and that negatively charged amino acid residues between positions 110 and 120 of 2a contribute to interaction with the 1a helicase-like domain but not to the intrinsic polymerase activity. Confocal fluorescence microscopy revealed that the BMV 1a and 2a colocalized to perinuclear region in human cells. However, no perinuclear spherule-like structures were detected in human cells by immunoelectron microscopy. Sequencing of the RNAs coimmunoprecipitated with RIG-I revealed that the 2a-synthesized short RNAs are derived from the message used to translate 2a. That is, 2a exhibits a strong cis preference for BMV RNA2. Strikingly, the 2a RNA products had initiation sequences (5'-GUAAA-3') identical to those from the 5' sequence of the BMV genomic RNA2 and RNA3. These results show that the BMV 2a polymerase does not require other BMV proteins to initiate RNA synthesis but that the 1a helicase domain, and likely helicase activity, can affect RNA synthesis by 2a.
Figures
References
-
- Adkins S, Kao C. 1998. Subgenomic RNA promoters dictate the mode of recognition by bromoviral RNA-dependent RNA polymerases. Virology 252:1–8 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
