

Dual transcriptional activities of SIX proteins define their roles in normal and ectopic eye development
- PMID: 22318629
- PMCID: PMC3274360
- DOI: 10.1242/dev.077255
Dual transcriptional activities of SIX proteins define their roles in normal and ectopic eye development
Abstract
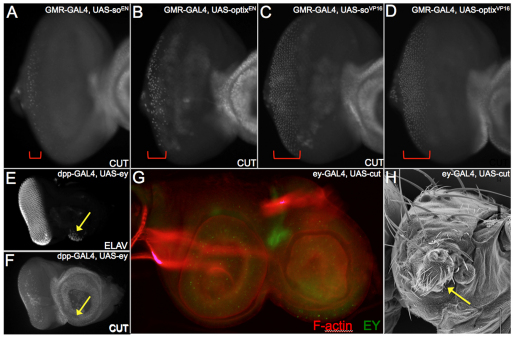
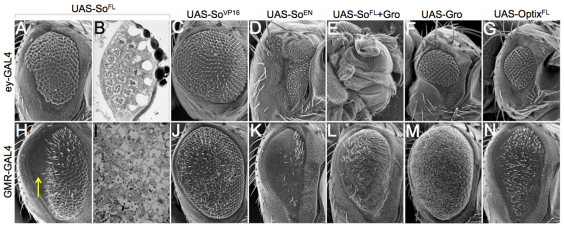
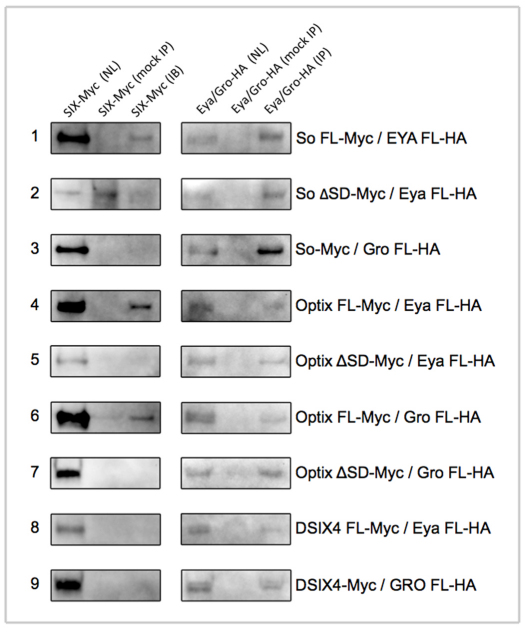
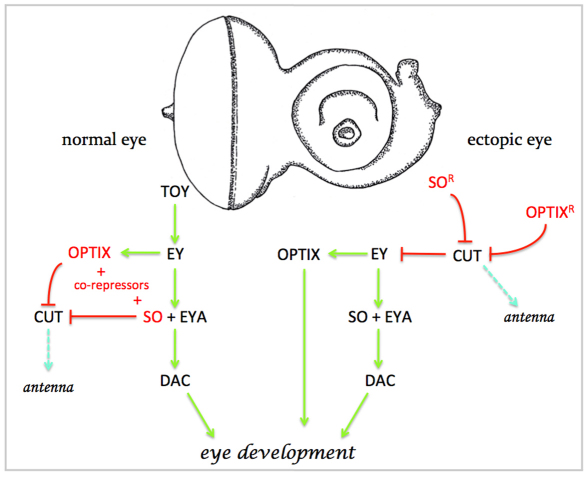
The SIX family of homeodomain-containing DNA-binding proteins play crucial roles in both Drosophila and vertebrate retinal specification. In flies, three such family members exist, but only two, Sine oculis (So) and Optix, are expressed and function within the eye. In vertebrates, the homologs of Optix (Six3 and Six6) and probably So (Six1 and Six2) are also required for proper eye formation. Depending upon the individual SIX protein and the specific developmental context, transcription of target genes can either be activated or repressed. These activities are thought to occur through physical interactions with the Eyes absent (Eya) co-activator and the Groucho (Gro) co-repressor, but the relative contribution that each complex makes to overall eye development is not well understood. Here, we attempt to address this issue by investigating the role that each complex plays in the induction of ectopic eyes in Drosophila. We fused the VP16 activation and Engrailed repressor domains to both So and Optix, and attempted to generate ectopic eyes with these chimeric proteins. Surprisingly, we find that So and Optix must initially function as transcriptional repressors to trigger the formation of ectopic eyes. Both factors appear to be required to repress the expression of non-retinal selector genes. We propose that during early phases of eye development, SIX proteins function, in part, to repress the transcription of non-retinal selector genes, thereby allowing induction of the retina to proceed. This model of repression-mediated induction of developmental programs could have implications beyond the eye and might be applicable to other systems.
Figures
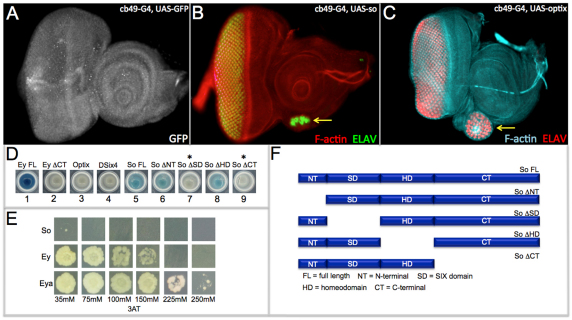
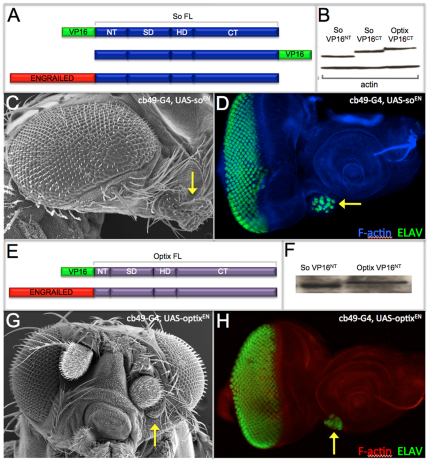
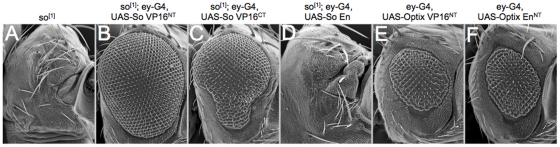
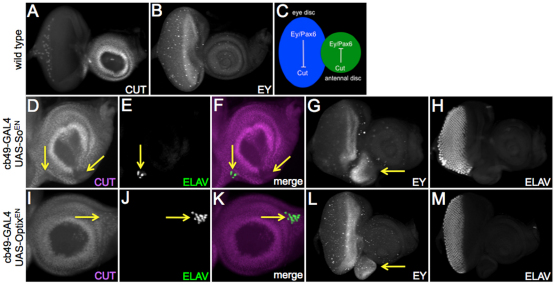
References
-
- Aldaz S., Morata G., Azpiazu N. (2003). The Pax-homeobox gene eyegone is involved in the subdivision of the thorax of Drosophila. Development 130, 4473–4482 - PubMed
-
- Altmann C. R., Chow R. L., Lang R. A., Hemmati-Brivanlou A. (1997). Lens induction by Pax-6 in Xenopus laevis. Dev. Biol. 185, 119–123 - PubMed
-
- Bischof J., Basler K. (2008). Recombinases and their use in gene activation, gene inactivation, and transgenesis. Methods Mol. Biol. 420, 175–195 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
