

Multilocus genetic analyses differentiate between widespread and spatially restricted cryptic species in a model ascidian
- PMID: 22319123
- PMCID: PMC3350675
- DOI: 10.1098/rspb.2011.2610
Multilocus genetic analyses differentiate between widespread and spatially restricted cryptic species in a model ascidian
Abstract
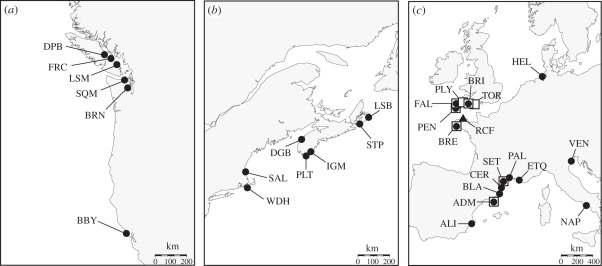
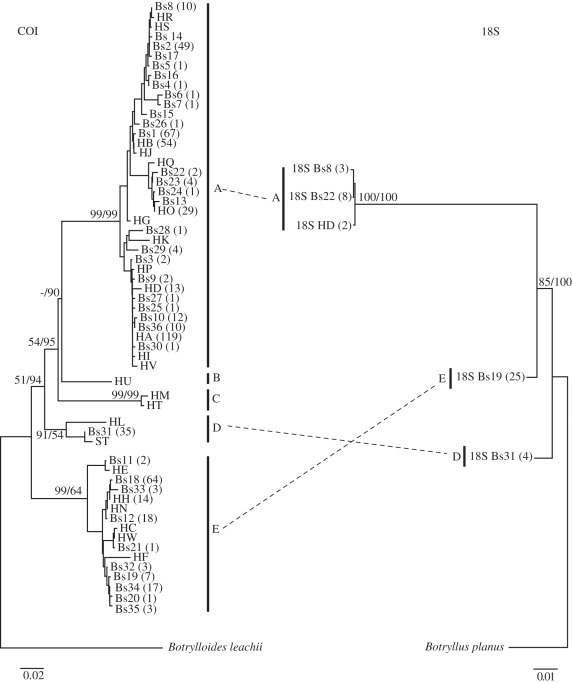
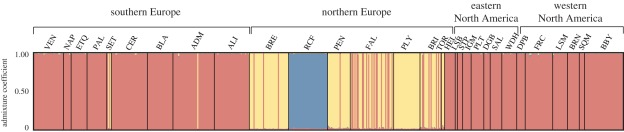
Elucidating the factors that shape species distributions has long been a fundamental goal in ecology and evolutionary biology. In spite of significant theoretical advancements, empirical studies of range limits have lagged behind. Specifically, little is known about how the attributes that allow species to expand their ranges and become widespread vary across phylogenies. Here, we studied the ascidian Botryllus schlosseri, a worldwide invasive species that is also characterized by marked genetic subdivision. Our study includes phylogenetic and population genetic data based on mitochondrial and nuclear genes, as well as polymorphic microsatellites for B. schlosseri colonies sampled from the southern and northern coasts of Europe and the eastern and western coasts of North America. We demonstrate that this well-known model organism comprises three highly divergent and probably reproductively isolated cryptic species (A, D and E), with two more (B and C) being suggested by data retrieved from GenBank. Among these, species A, recovered in all of the surveyed regions, is by far the most common and widespread. By contrast, species B-E, occurring mostly in sites from northern Europe, are considerably more geographically restricted. These findings, along with inferences made on transport opportunity, suggest that divergent evolutionary histories promoted differences in invasive potential between B. schlosseri sibling species, indicating that attributes that facilitate dramatic shifts in range limits can evolve more easily and frequently than previously thought. We propose environmental disturbance as a selective force that could have shaped the evolution of invasiveness in the B. schlosseri complex.
Figures
References
-
- Elton C. S. 1930. Animal ecology and evolution. Oxford, UK: The Clarendon Press
-
- Sexton J. P., McIntyre P. J., Angert A. L., Rice K. J. 2009. Evolution and ecology of species range limits. Annu. Rev. Ecol. Evol. Syst. 40, 415–43610.1146/annurev.ecolsys.110308.120317 (doi:10.1146/annurev.ecolsys.110308.120317) - DOI - DOI
-
- Keitt T. H., Lewis M. A., Holt R. D. 2001. Allee effects, invasion pinning, and species' borders. Am. Nat. 157, 203–21610.1086/318633 (doi:10.1086/318633) - DOI - DOI - PubMed
-
- Pulliam H. R. 2000. On the relationship between niche and distribution. Ecol. Lett. 3, 349–36110.1046/j.1461-0248.2000.00143.x (doi:10.1046/j.1461-0248.2000.00143.x) - DOI - DOI
-
- Kirkpatrick M., Barton N. H. 1997. Evolution of a species' range. Am. Nat. 150, 1–2310.1086/286054 (doi:10.1086/286054) - DOI - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
