

Pericellular pH homeostasis is a primary function of the Warburg effect: inversion of metabolic systems to control lactate steady state in tumor cells
- PMID: 22320183
- PMCID: PMC3294166
- DOI: 10.1111/j.1349-7006.2012.02206.x
Pericellular pH homeostasis is a primary function of the Warburg effect: inversion of metabolic systems to control lactate steady state in tumor cells
Abstract
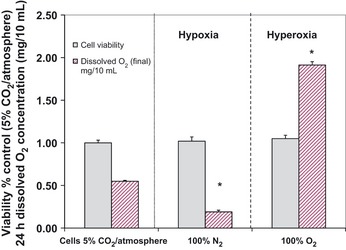
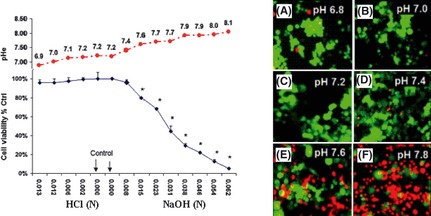
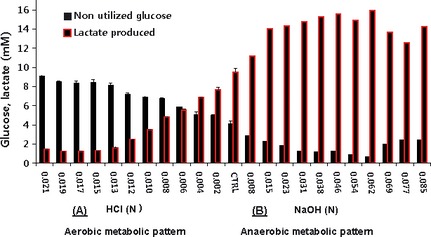
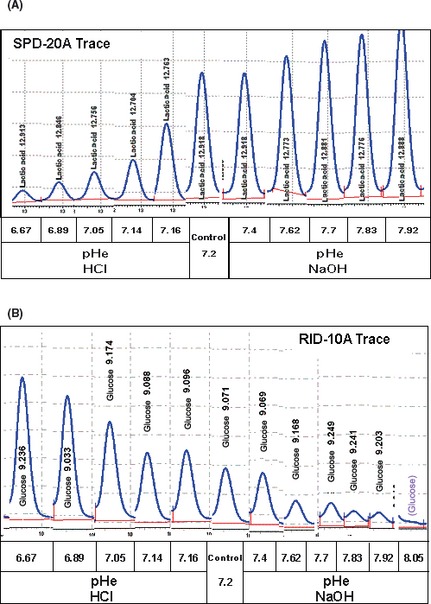
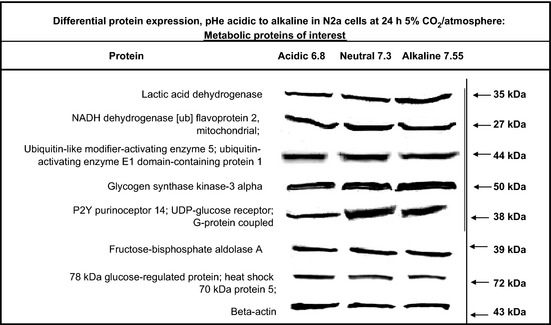
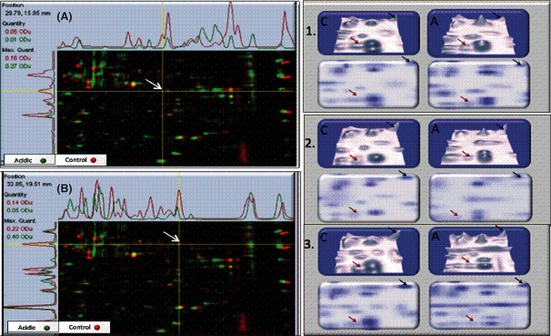
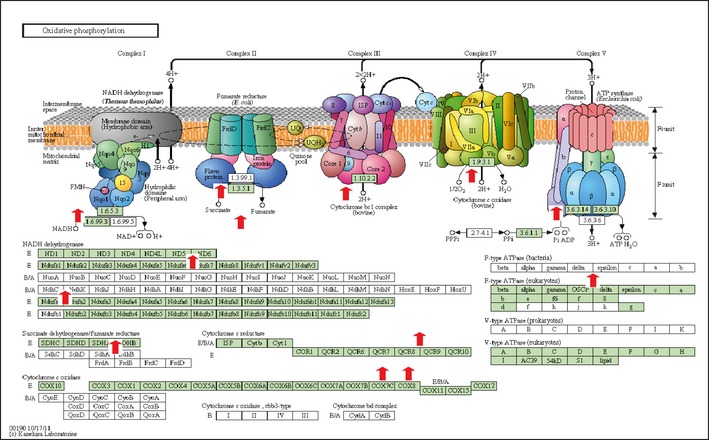
The Warburg effect describes a heightened propensity of tumor cells to produce lactic acid in the presence or absence of O(2) . A generally held notion is that the Warburg effect is related to energy. Using whole-genome, proteomic MALDI-TOF-MS and metabolite analysis, we investigated the Warburg effect in malignant neuroblastoma N2a cells. The findings show that the Warburg effect serves a functional role in regulating acidic pericellular pH (pHe), which is mediated by metabolic inversion or a fluctuating dominance between glycolytic-rate substrate level phosphorylation (SLP) and mitochondrial (mt) oxidative phosphorylation (OXPHOS) to control lactic acid production. The results also show that an alkaline pHe caused an elevation in SLP/OXPHOS ratio (approximately 98% SLP/OXPHOS); while the ratio was approximately 56% at neutral pHe and approximately 93% in acidic pHe. Acidic pHe paralleled greater expression of mitochondrial biogenesis and OXPHOS genes, such as complex III-V (Uqcr10, Atp5 and Cox7c), mt Fmc1, Romo1, Tmem 173, Tomm6, aldehyde dehydrogenase, mt Sod2 mt biogenesis component PPAR-γ co-activator 1 adjunct to loss of mt fission (Mff). Moreover, acidic pHe corresponded to metabolic efficiency evidenced by a rise in mTOR nutrient sensor GβL, its downstream target (Eif4ebp1), insulin modulators (Trib3 and Fetub) and loss of catabolic (Hadhb, Bdh1 and Pygl)/glycolytic processes (aldolase C, pyruvate kinase, Nampt and aldose-reductase). In contrast, alkaline pHe initiated loss of mitofusin 2, complex II-IV (Sdhaf1, Uqcrq, Cox4i2 and Aldh1l2), aconitase, mitochondrial carrier triple repeat 1 and mt biosynthetic (Coq2, Coq5 and Coq9). In conclusion, the Warburg effect might serve as a negative feedback loop that regulates the pHe toward a broad acidic range by altering lactic acid production through inversion of metabolic systems. These effects were independent of changes in O(2) concentration or glucose supply.
© 2012 Japanese Cancer Association.
Figures

References
-
- Champion L, Brain E, Giraudet AL et al Breast cancer recurrence diagnosis suspected on tumor marker rising: value of whole‐body 18FDG‐PET/CT imaging and impact on patient management. Cancer 2011; 117: 1621–9. - PubMed
-
- Mentis AF, Kararizou E. Metabolism and cancer: an up‐to‐date review of a mutual connection. Asian Pac J Cancer Prev 2010; 11: 1437–44. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
