

Biophysics of mitosis
- PMID: 22321376
- PMCID: PMC4433171
- DOI: 10.1017/S0033583512000017
Biophysics of mitosis
Abstract
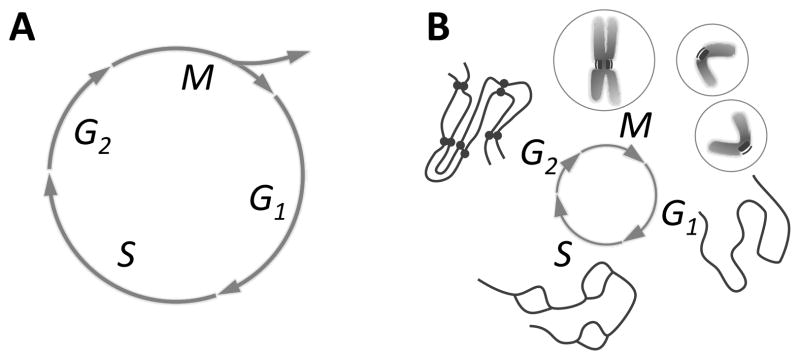
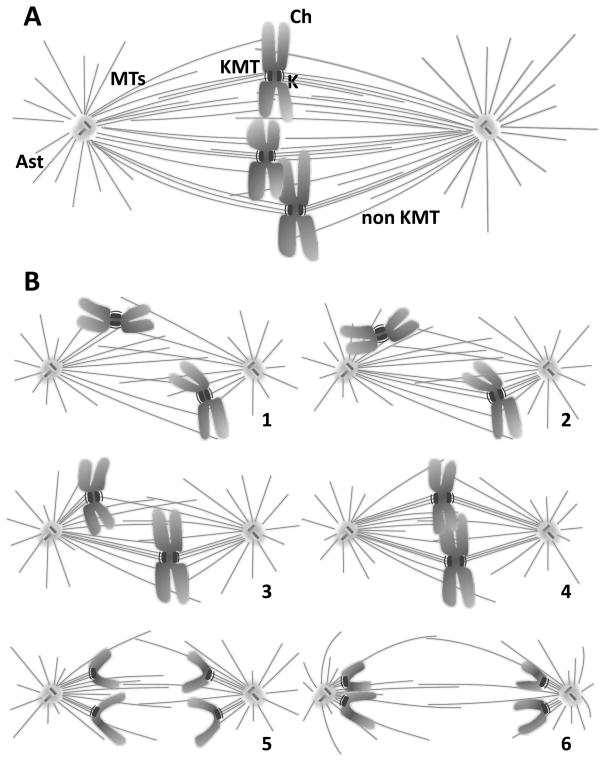
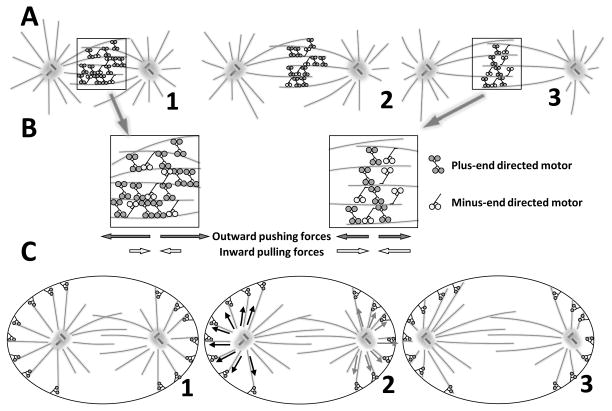
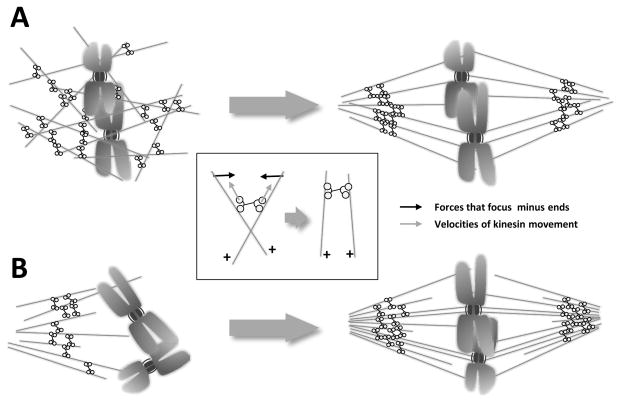
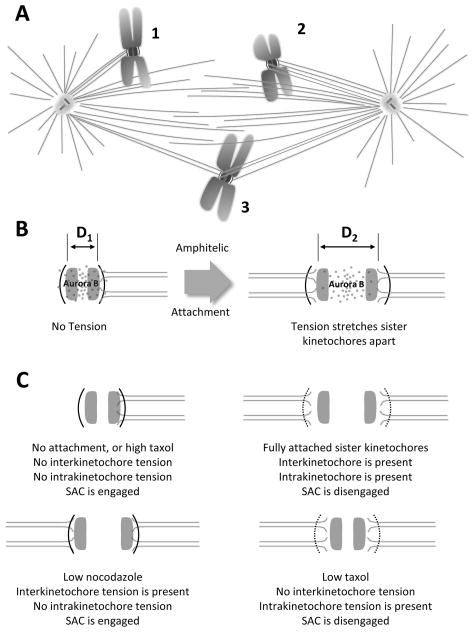
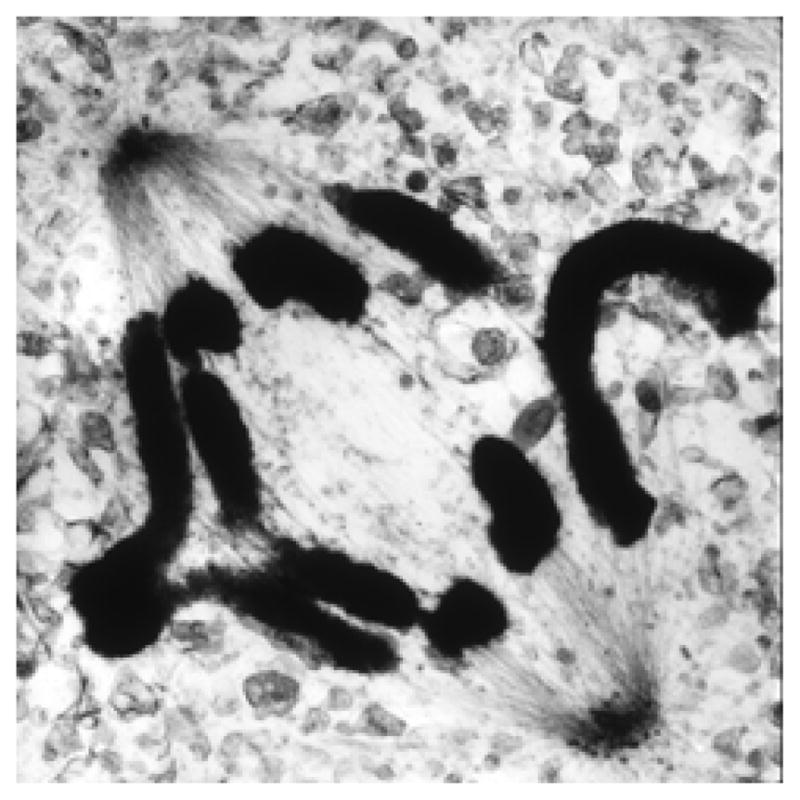
Mitosis is the process by which eukaryotic cells organize and segregate their chromosomes in preparation for cell division. It is accomplished by a cellular machine composed largely of microtubules (MTs) and their associated proteins. This article reviews literature on mitosis from a biophysical point of view, drawing attention to the assembly and motility processes required to do this complex job with precision. Work from both the recent and the older literature is integrated into a description of relevant biological events and the experiments that probe their mechanisms. Theoretical work on specific subprocesses is also reviewed. Our goal is to provide a document that will expose biophysicists to the fascination of this quite amazing process and provide them with a good background from which they can pursue their own research interests in the subject.
Figures

References
-
- AIST JR, BAYLES CJ, TAO W, BERNS MW. Direct experimental evidence for the existence, structural basis and function of astral forces during anaphase B in vivo. J Cell Sci. 1991;100 (Pt 2):279–288. - PubMed
-
- ALTAN-BONNET N, SOUGRAT R, LIPPINCOTT-SCHWARTZ J. Molecular basis for Golgi maintenance and biogenesis. Curr Opin Cell Biol. 2004;16(4):364–372. - PubMed
-
- ANTONIO C, FERBY I, WILHELM H, JONES M, KARSENTI E, NEBREDA AR, VERNOS I. Xkid, a chromokinesin required for chromosome alignment on the metaphase plate. Cell. 2000;102(4):425–435. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
