

F-box protein FBXL2 targets cyclin D2 for ubiquitination and degradation to inhibit leukemic cell proliferation
- PMID: 22323446
- PMCID: PMC3321873
- DOI: 10.1182/blood-2011-06-358911
F-box protein FBXL2 targets cyclin D2 for ubiquitination and degradation to inhibit leukemic cell proliferation
Abstract
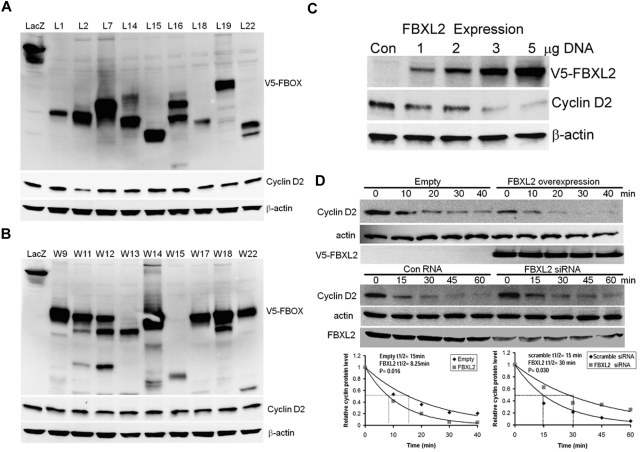
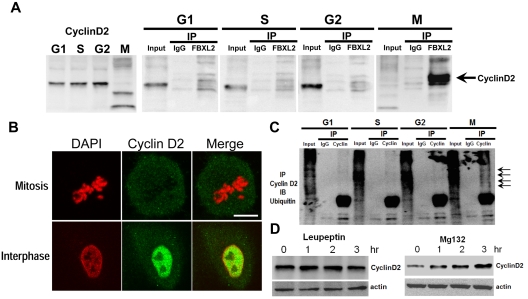
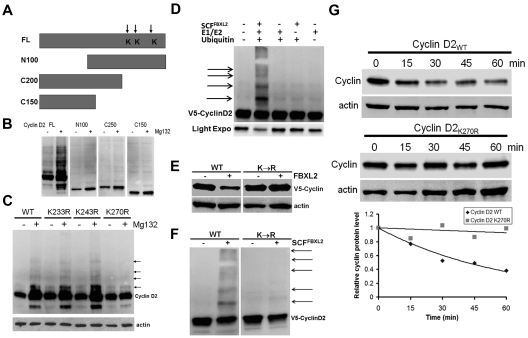
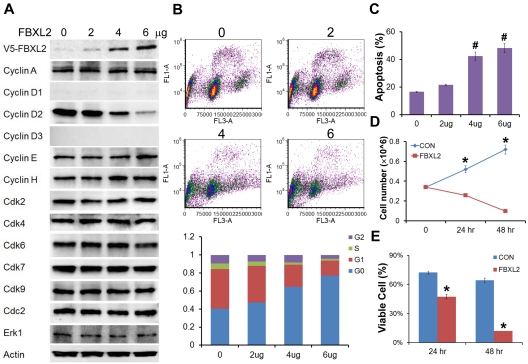
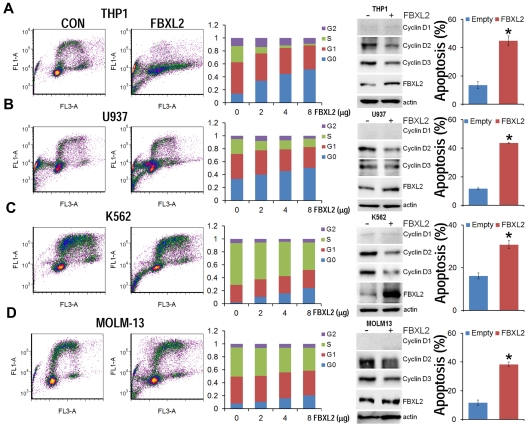
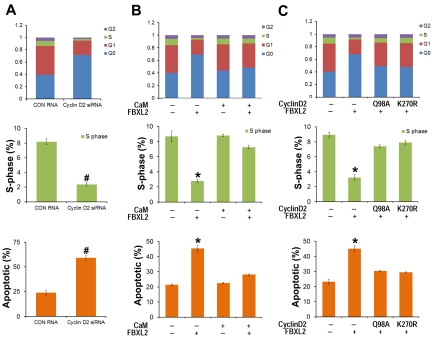
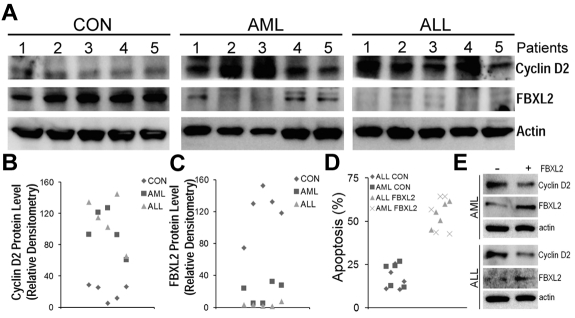
Hematologic maligancies exhibit a growth advantage by up-regulation of components within the molecular apparatus involved in cell-cycle progression. The SCF (Skip-Cullin1-F-box protein) E3 ligase family provides homeostatic feedback control of cell division by mediating ubiquitination and degradation of cell-cycle proteins. By screening several previously undescribed E3 ligase components, we describe the behavior of a relatively new SCF subunit, termed FBXL2, that ubiquitinates and destabilizes cyclin D2 protein leading to G(0) phase arrest and apoptosis in leukemic and B-lymphoblastoid cell lines. FBXL2 expression was strongly suppressed, and yet cyclin D2 protein levels were robustly expressed in acute myelogenous leukemia (AML) and acute lymphoblastic leukemia (ALL) patient samples. Depletion of endogenous FBXL2 stabilized cyclin D2 levels, whereas ectopically expressed FBXL2 decreased cyclin D2 lifespan. FBXL2 did not bind a phosphodegron within its substrate, which is typical of other F-box proteins, but uniquely targeted a calmodulin-binding signature within cyclin D2 to facilitate its polyubiquitination. Calmodulin competes with the F-box protein for access to this motif where it bound and protected cyclin D2 from FBXL2. Calmodulin reversed FBXL2-induced G(0) phase arrest and attenuated FBXL2-induced apoptosis of lymphoblastoid cells. These results suggest an antiproliferative effect of SCF(FBXL2) in lymphoproliferative malignancies.
Figures
References
-
- Buchakjian MR, Kornbluth S. The engine driving the ship: metabolic steering of cell proliferation and death. Nat Rev Mol Cell Biol. 2010;11(10):715–727. - PubMed
-
- Siebert R, Willers CP, Opalka B. Role of the cyclin-dependent kinase 4 and 6 inhibitor gene family p15, p16, p18, and p19 in leukemia and lymphoma. Leuk Lymphoma. 1996;23(5-6):505–520. - PubMed
-
- Lew DJ, Dulic V, Reed SI. Isolation of three novel human cyclins by rescue of G1 cyclin (Cln) function in yeast. Cell. 1991;66(6):1197–1206. - PubMed
-
- Gautschi O, Ratschiller D, Gugger M, Betticher DC, Heighway J. Cyclin D1 in non-small cell lung cancer: a key driver of malignant transformation. Lung Cancer. 2007;55(1):1–14. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
