

Remembering to learn: independent place and journey coding mechanisms contribute to memory transfer
- PMID: 22323731
- PMCID: PMC3567459
- DOI: 10.1523/JNEUROSCI.3998-11.2012
Remembering to learn: independent place and journey coding mechanisms contribute to memory transfer
Abstract
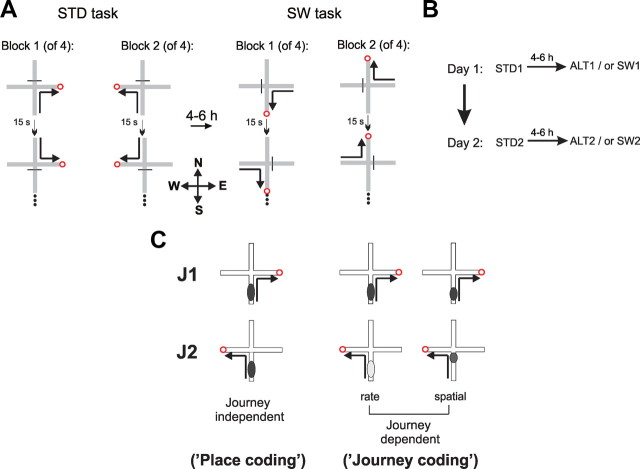
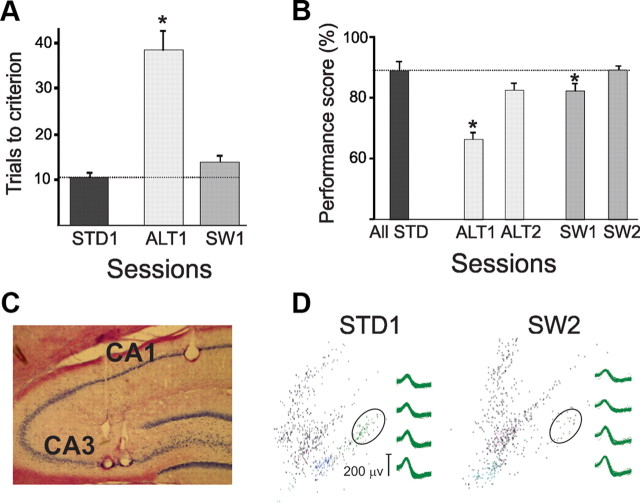
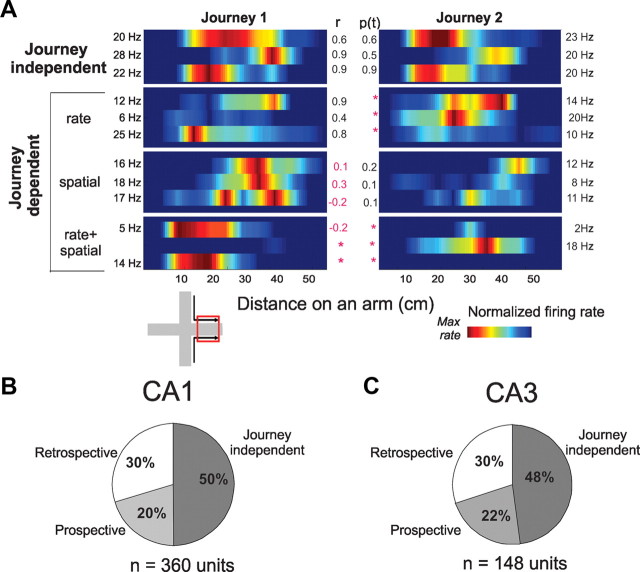
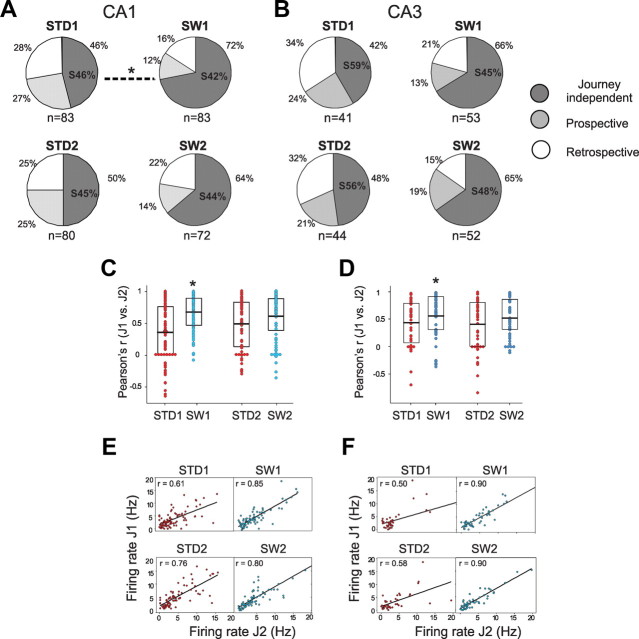
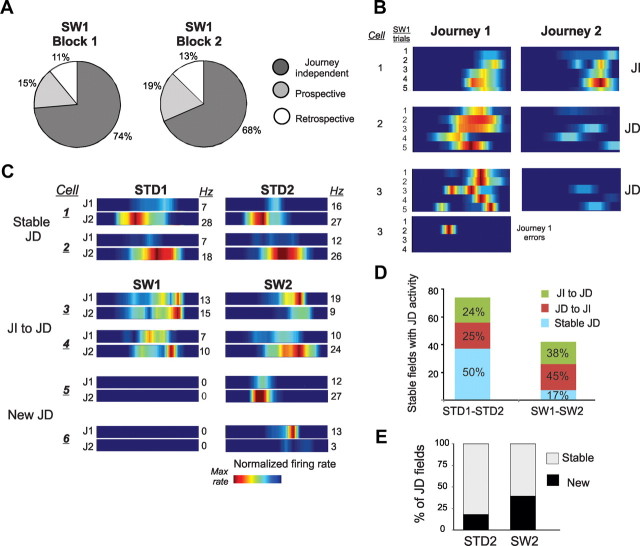
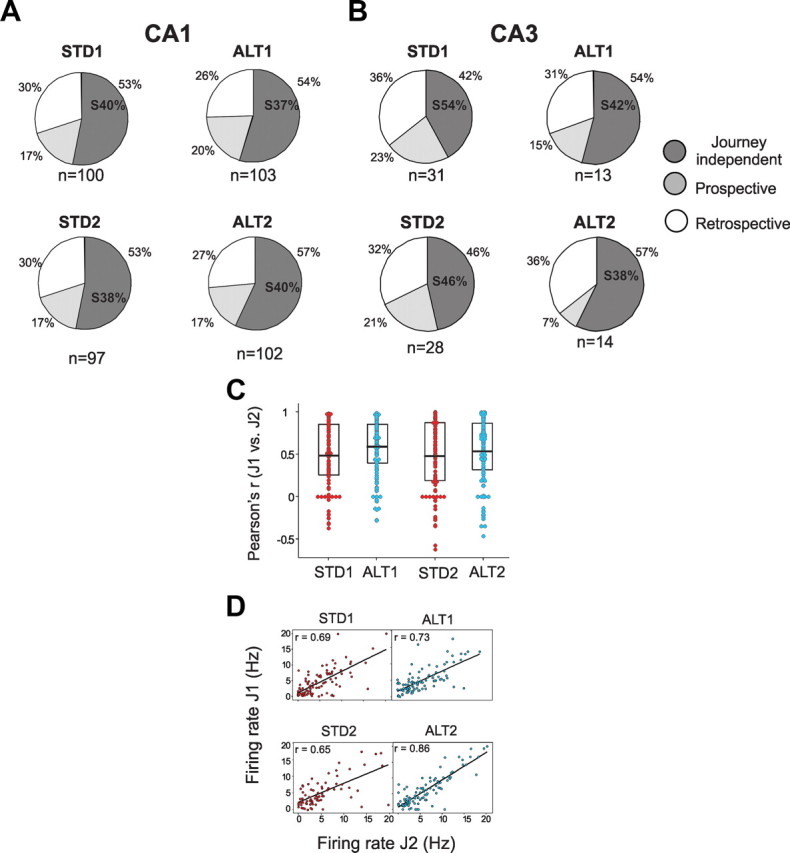
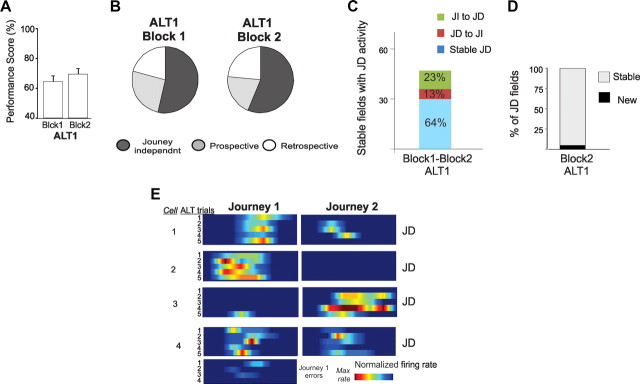
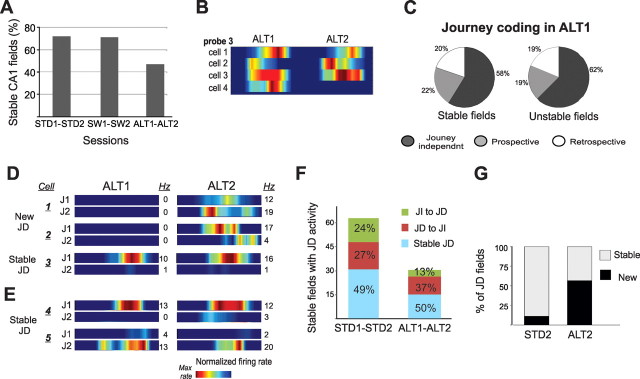
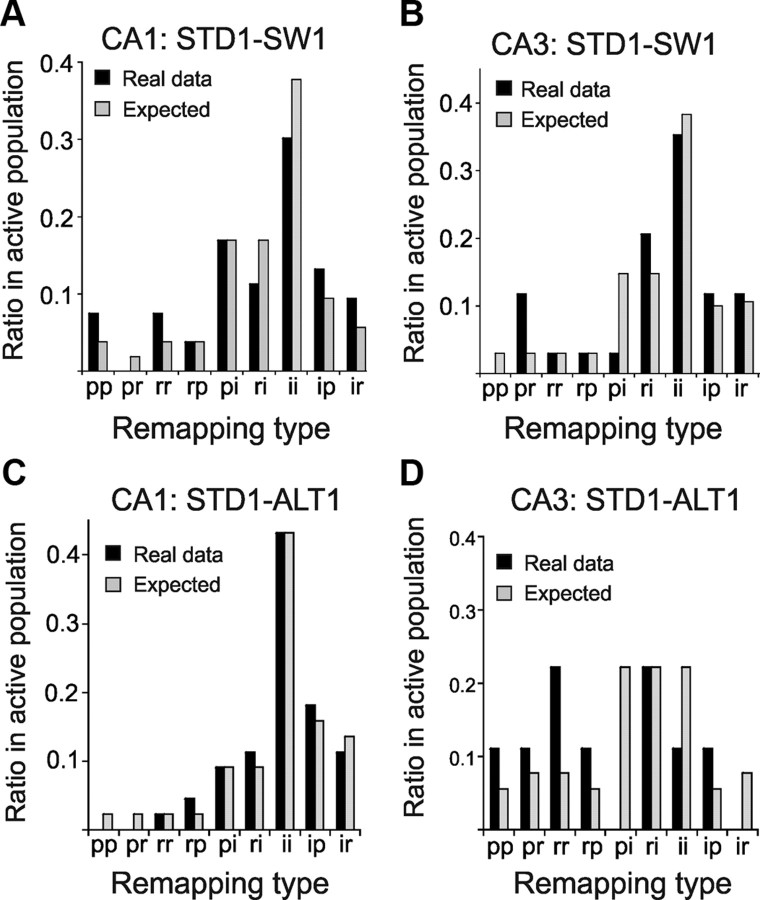
The neural mechanisms that integrate new episodes with established memories are unknown. When rats explore an environment, CA1 cells fire in place fields that indicate locations. In goal-directed spatial memory tasks, some place fields differentiate behavioral histories ("journey-dependent" place fields) while others do not ("journey-independent" place fields). To investigate how these signals inform learning and memory for new and familiar episodes, we recorded CA1 and CA3 activity in rats trained to perform a "standard" spatial memory task in a plus maze and in two new task variants. A "switch" task exchanged the start and goal locations in the same environment; an "altered environment" task contained unfamiliar local and distal cues. In the switch task, performance was mildly impaired, new firing maps were stable, but the proportion and stability of journey-dependent place fields declined. In the altered environment, overall performance was strongly impaired, new firing maps were unstable, and stable proportions of journey-dependent place fields were maintained. In both tasks, memory errors were accompanied by a decline in journey codes. The different dynamics of place and journey coding suggest that they reflect separate mechanisms and contribute to distinct memory computations. Stable place fields may represent familiar relationships among environmental features that are required for consistent memory performance. Journey-dependent activity may correspond with goal-directed behavioral sequences that reflect expectancies that generalize across environments. The complementary signals could help link current events with established memories, so that familiarity with either a behavioral strategy or an environment can inform goal-directed learning.
Figures
References
-
- Brun VH, Otnass MK, Molden S, Steffenach HA, Witter MP, Moser MB, Moser EI. Place cells and place recognition maintained by direct entorhinal-hippocampal circuitry. Science. 2002;296:2243–2246. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous