

Rabbit haemorrhagic disease (RHD) and rabbit haemorrhagic disease virus (RHDV): a review
- PMID: 22325049
- PMCID: PMC3331820
- DOI: 10.1186/1297-9716-43-12
Rabbit haemorrhagic disease (RHD) and rabbit haemorrhagic disease virus (RHDV): a review
Abstract
Rabbit haemorrhagic disease virus (RHDV) is a calicivirus of the genus Lagovirus that causes rabbit haemorrhagic disease (RHD) in adult European rabbits (Oryctolagus cuniculus). First described in China in 1984, the virus rapidly spread worldwide and is nowadays considered as endemic in several countries. In Australia and New Zealand where rabbits are pests, RHDV was purposely introduced for rabbit biocontrol. Factors that may have precipitated RHD emergence remain unclear, but non-pathogenic strains seem to pre-date the appearance of the pathogenic strains suggesting a key role for the comprehension of the virus origins. All pathogenic strains are classified within one single serotype, but two subtypes are recognised, RHDV and RHDVa. RHD causes high mortality in both domestic and wild adult animals, with individuals succumbing between 48-72 h post-infection. No other species has been reported to be fatally susceptible to RHD. The disease is characterised by acute necrotising hepatitis, but haemorrhages may also be found in other organs, in particular the lungs, heart, and kidneys due to disseminated intravascular coagulation. Resistance to the disease might be explained in part by genetically determined absence or weak expression of attachment factors, but humoral immunity is also important. Disease control in rabbitries relies mainly on vaccination and biosecurity measures. Such measures are difficult to be implemented in wild populations. More recent research has indicated that RHDV might be used as a molecular tool for therapeutic applications. Although the study of RHDV and RHD has been hampered by the lack of an appropriate cell culture system for the virus, several aspects of the replication, epizootology, epidemiology and evolution have been disclosed. This review provides a broad coverage and description of the current knowledge on the disease and the virus.
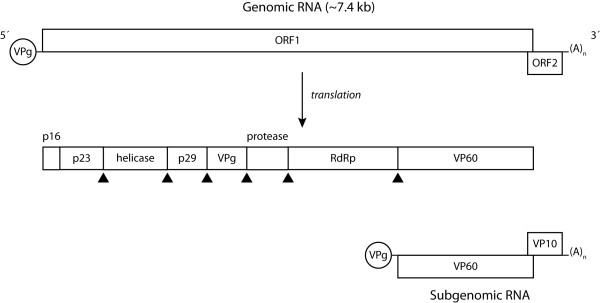
Figures



References
-
- Liu SJ, Xue HP, Pu BQ, Qian NH. A new viral disease in rabbit. Anim Husb Vet Med. 1984;16:253–255.
-
- Xu WY. Viral haemorrhagic disease of rabbits in the People's Republic of China: epidemiology and virus characterisation. Rev Sci Tech. 1991;10:393–408. - PubMed
-
- Park NY, Chong CY, Kim JH, Cho SM, Cha YH, Jung BT, Kim DS, Yoon JB. An outbreak of viral haemorrhagic pneumonia (tentative name) of rabbits in Korea. J Korean Vet Med Assoc. 1987;23:603–610.
-
- Cancellotti FM, Renzi M. Epidemiology and current situation of viral haemorrhagic disease of rabbits and the European brown hare syndrome in Italy. Rev Sci Tech. 1991;10:409–422. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
