

Hypermutation of ApoB mRNA by rat APOBEC-1 overexpression mimics APOBEC-3 hypermutation
- PMID: 22326345
- PMCID: PMC3311742
- DOI: 10.1016/j.jmb.2012.02.005
Hypermutation of ApoB mRNA by rat APOBEC-1 overexpression mimics APOBEC-3 hypermutation
Abstract
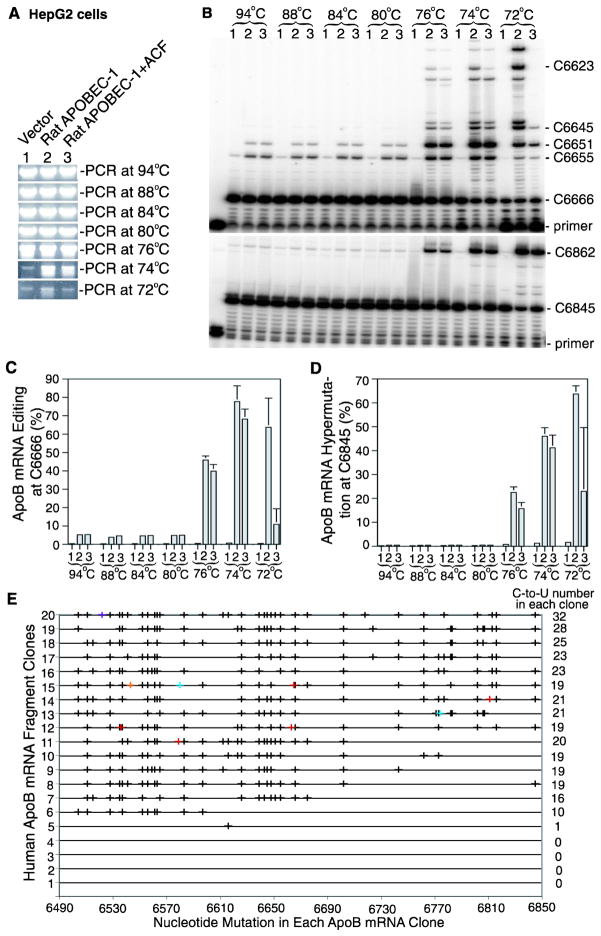
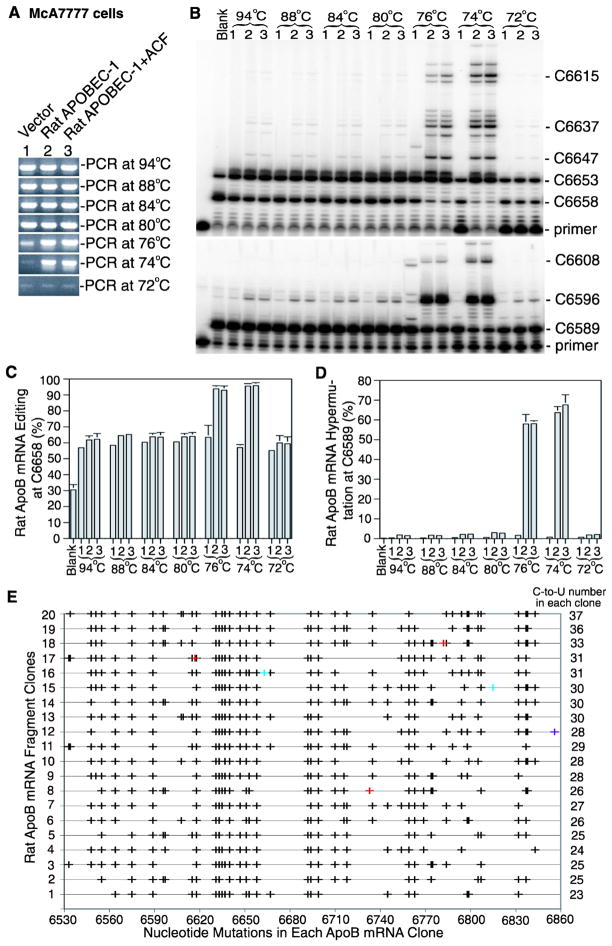
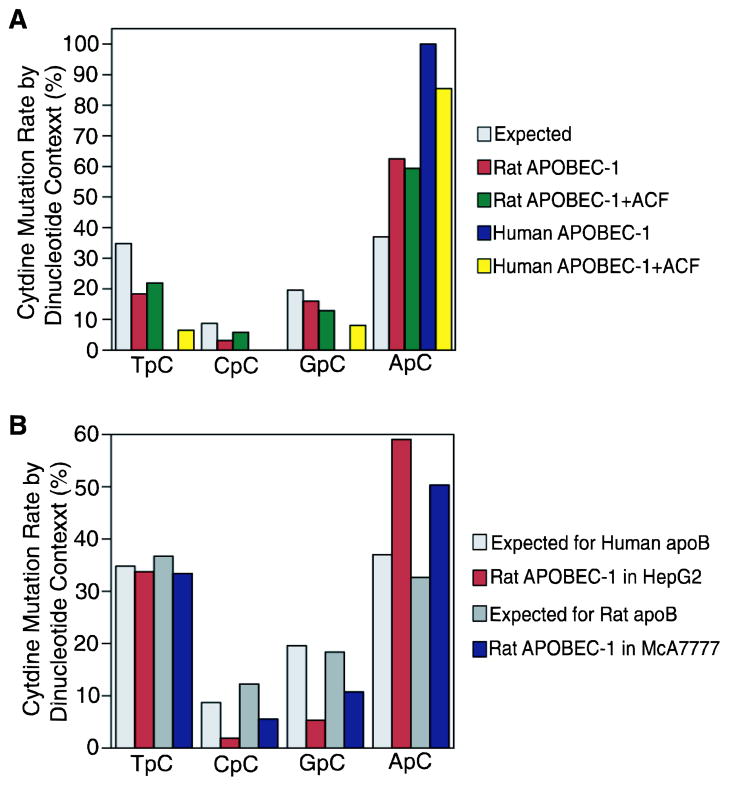
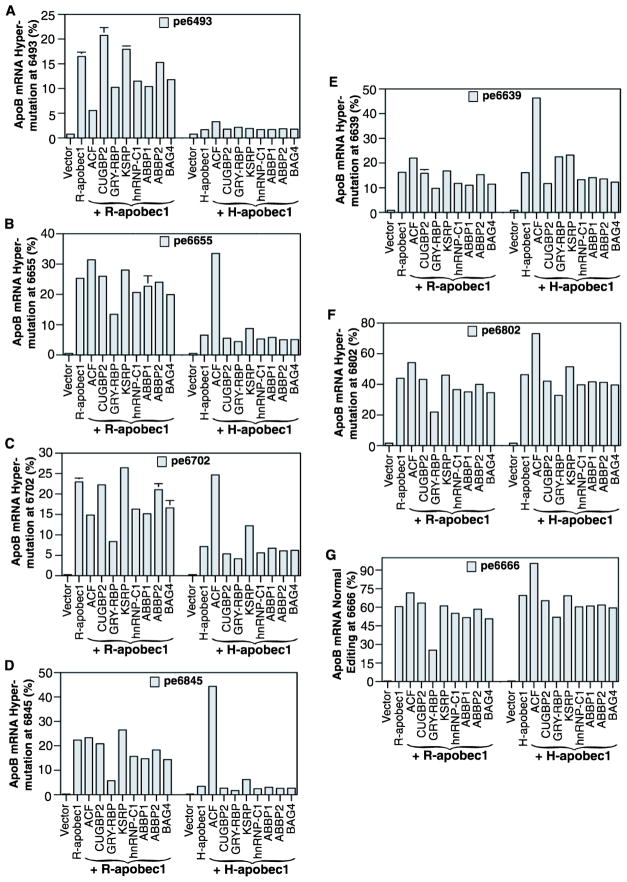
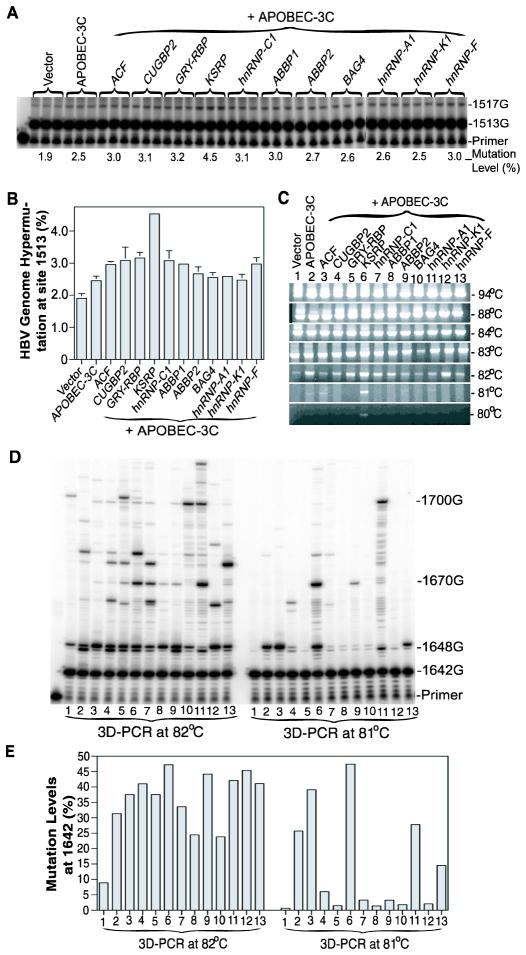
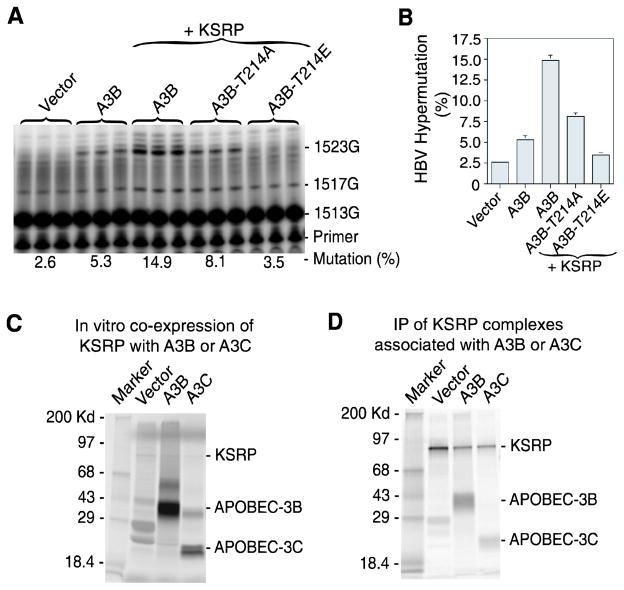
APOBEC-3 proteins induce C-to-U hypermutations in the viral genome of various viruses and have broad antiviral activity. Generally, only a small proportion of viral genomes (<10(-)(2)) are hypermutated by APOBEC-3s, but often many cytidines (up to 40%) are converted into uridine. The mechanism of this unique selective hypermutation remains unknown. We found that rat APOBEC-1 overexpression had a hypermutation pattern similar to that of APOBEC-3s on its substrate apolipoprotein B (apoB) mRNA. Transient plasmid transfection of rat APOBEC-1 resulted in 0.4% and 1.8% hypermutations with apoB mRNA in HepG2 and McA7777 cells, respectively. The low frequency of hypermutated apoB mRNA targets was enriched by differential DNA denaturation PCR at 72-76 °C, with hypermutation levels increasing up to 67%. Up to 69.6% of cytidines in HepG2 and up to 75.5% of cytidines in McA7777 cells were converted into uridines in the hypermutated apoB mRNA. When rat APOBEC-1 was overexpressed by adenovirus, the hypermutation frequency of apoB mRNA increased from 0.4% to ∼20% and was readily detected by regular PCR. However, this higher expression efficiency only increased the frequency of hypermutation, not the number of affected cytidines in hypermutated targets. Rat APOBEC-1 hypermutation was modulated by cofactors and eliminated by an E181Q mutation, indicating the role of cofactors in hypermutation. The finding of an APOBEC-3 hypermutation pattern with rat APOBEC-1 suggests that cofactors could also be involved in APOBEC-3 hypermutation. Using hepatitis B virus hypermutation, we found that KSRP increased APOBEC-3C and APOBEC-3B hypermutation. These data show that, like rat APOBEC-1 hypermutation, cellular factors may play a regulatory role in APOBEC-3 hypermutation.
Published by Elsevier Ltd.
Figures

References
-
- Sheehy AM, Gaddis NC, Choi JD, Malim MH. Isolation of a human gene that inhibits HIV-1 infection and is suppressed by the viral Vif protein. Nature. 2002;418:646–650. - PubMed
-
- Yu Q, Konig R, Pillai S, Chiles K, Kearney M, Palmer S, Richman D, Coffin JM, Landau NR. Single-strand specificity of APOBEC3G accounts for minus-strand deamination of the HIV genome. Nat Struct Mol Biol. 2004;11:435–442. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
