

Frequent alterations and epigenetic silencing of differentiation pathway genes in structurally rearranged liposarcomas
- PMID: 22328974
- PMCID: PMC3274776
- DOI: 10.1158/2159-8290.CD-11-0181
Frequent alterations and epigenetic silencing of differentiation pathway genes in structurally rearranged liposarcomas
Abstract
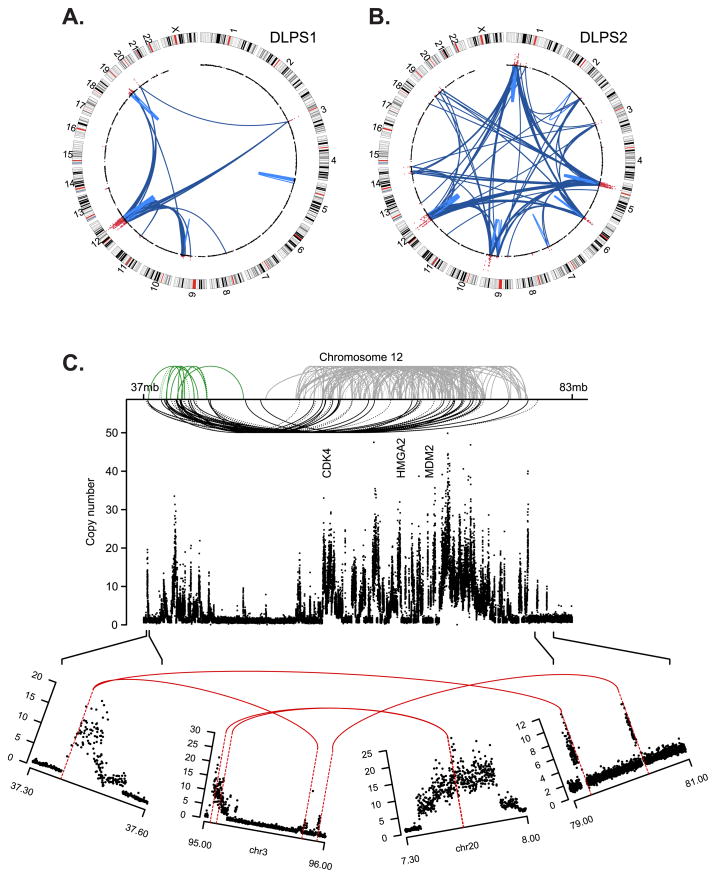
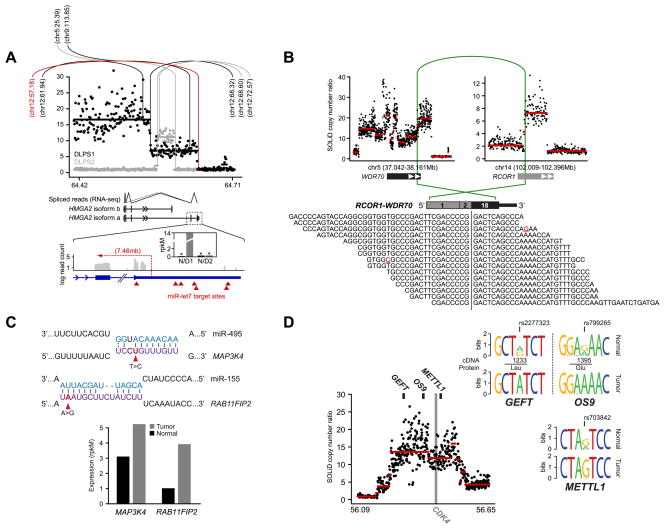
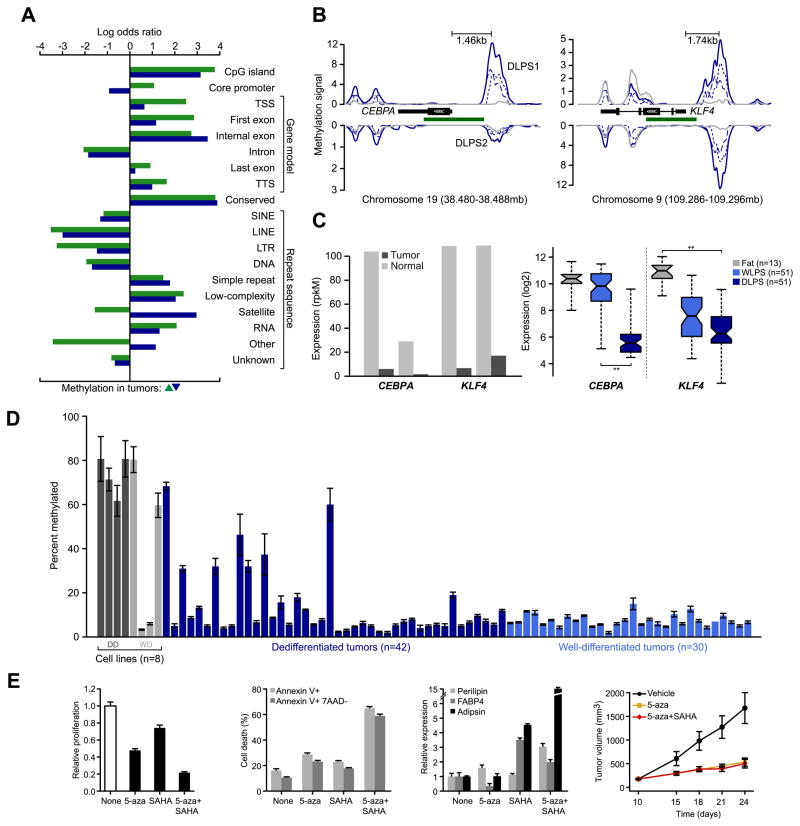
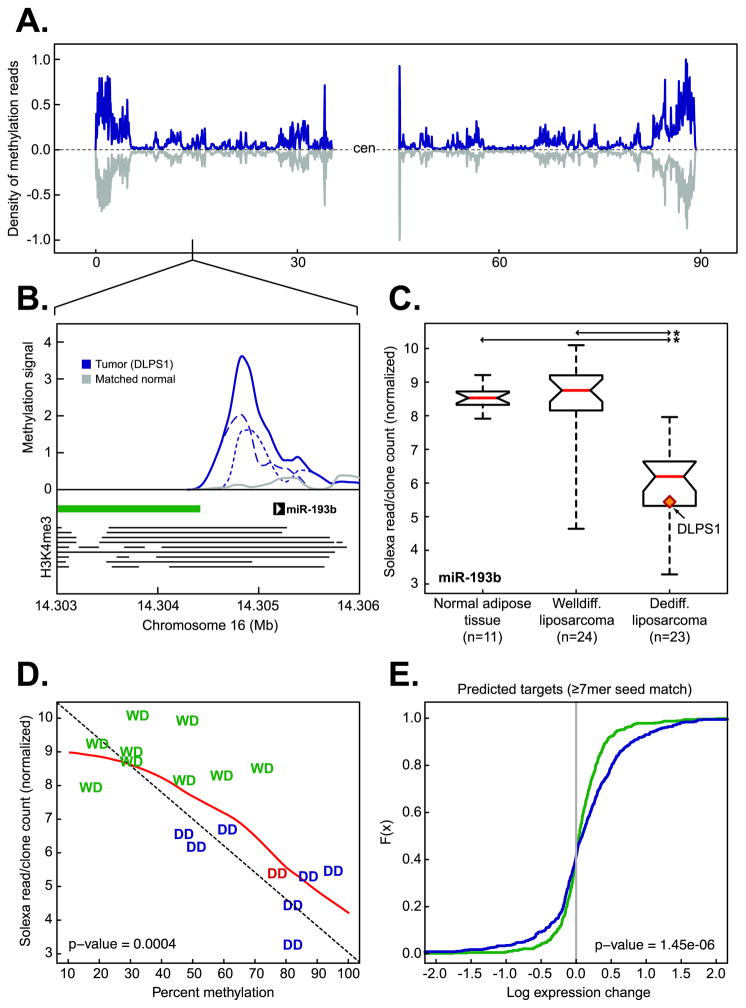
We explored diverse alterations contributing to liposarcomagenesis by sequencing the genome, exome, transcriptome, and cytosine methylome of a primary and recurrent dedifferentiated liposarcoma (DLPS) from distinct chemotherapy/radiotherapy-naïve patients. The liposarcoma genomes had complex structural rearrangements, but in different patterns, and with varied effects on the structure and expression of affected genes. While the point mutation rate was modest, integrative analyses and additional screening identified somatic mutations in HDAC1 in 8.3% of DLPS. Liposarcoma methylomes revealed alterations in differentiation pathway genes, including CEBPA methylation in 24% of DLPS. Treatment with demethylating agents, which restored CEBPA expression in DLPS cells, was anti-proliferative and pro-apoptotic in vitro and reduced tumor growth in vivo. Both genetic and epigenetic abnormalities established a role for small RNAs in liposarcomagenesis, typified by methylation-induced silencing of microRNA-193b in DLPS but not its well-differentiated counterpart. These findings reveal an unanticipated role for epigenetic abnormalities in DLPS tumors and suggest demethylating agents as potential therapeutics.
Significance: Multimodality sequence analysis of DLPS revealed recurrent mutations and epigenetic abnormalities critical to liposarcomagenesis and to the suppression of adipocyte differentiation. Pharmacologic inhibition of DNA methylation promoted apoptosis and differentiated DLPS cells in vitro and inhibited tumor growth in vivo, providing a rationale for investigating methylation inhibitors in this disease.
Keywords: DNA methylation; Dedifferentiated liposarcoma; adipocyte differentiation; histone deacetylase; microRNA.
Conflict of interest statement
Figures
Comment in
-
Genomic investigation of dedifferentiated liposarcoma suggests a role for therapeutic targeting of the tumor epigenome.Cancer Discov. 2011 Dec;1(7):555-6. doi: 10.1158/2159-8290.CD-11-0291. Cancer Discov. 2011. PMID: 22586679
References
-
- Fletcher C, Unni K, Mertens F. Pathology and genetics of tumors of soft tissue and bone. International Agency for Research on Cancer Press; Lyon: 2002.
-
- Pedeutour F, Forus A, Coindre JM, Berner JM, Nicolo G, Michiels JF, et al. Structure of the supernumerary ring and giant rod chromosomes in adipose tissue tumors. Genes Chromosomes Cancer. 1999;24:30–41. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
