

Calcium/calmodulin stimulates the autophosphorylation of elongation factor 2 kinase on Thr-348 and Ser-500 to regulate its activity and calcium dependence
- PMID: 22329831
- PMCID: PMC3401519
- DOI: 10.1021/bi201788e
Calcium/calmodulin stimulates the autophosphorylation of elongation factor 2 kinase on Thr-348 and Ser-500 to regulate its activity and calcium dependence
Abstract
Eukaryotic elongation factor 2 kinase (eEF-2K) is an atypical protein kinase regulated by Ca(2+) and calmodulin (CaM). Its only known substrate is eukaryotic elongation factor 2 (eEF-2), whose phosphorylation by eEF-2K impedes global protein synthesis. To date, the mechanism of eEF-2K autophosphorylation has not been fully elucidated. To investigate the mechanism of autophosphorylation, human eEF-2K was coexpressed with λ-phosphatase and purified from bacteria in a three-step protocol using a CaM affinity column. Purified eEF-2K was induced to autophosphorylate by incubation with Ca(2+)/CaM in the presence of MgATP. Analyzing tryptic or chymotryptic peptides by mass spectrometry monitored the autophosphorylation over 0-180 min. The following five major autophosphorylation sites were identified: Thr-348, Thr-353, Ser-445, Ser-474, and Ser-500. In the presence of Ca(2+)/CaM, robust phosphorylation of Thr-348 occurs within seconds of addition of MgATP. Mutagenesis studies suggest that phosphorylation of Thr-348 is required for substrate (eEF-2 or a peptide substrate) phosphorylation, but not self-phosphorylation. Phosphorylation of Ser-500 lags behind the phosphorylation of Thr-348 and is associated with the Ca(2+)-independent activity of eEF-2K. Mutation of Ser-500 to Asp, but not Ala, renders eEF-2K Ca(2+)-independent. Surprisingly, this Ca(2+)-independent activity requires the presence of CaM.
Figures



 ). (B) Sequence coverage of the autophosphorylated enzyme (3 h incubation with CaM/Ca2+ /MgATP) is ~ 86%, indicated by the residues in red (
). (B) Sequence coverage of the autophosphorylated enzyme (3 h incubation with CaM/Ca2+ /MgATP) is ~ 86%, indicated by the residues in red ( ). Both samples were resolved by SDS-PAGE, subjected to tryptic and chymotryptic in-gel digestion and the peptide digests then used for mass spectrometry analysis as described under ‘Supplementary Experimental Procedures’.
). Both samples were resolved by SDS-PAGE, subjected to tryptic and chymotryptic in-gel digestion and the peptide digests then used for mass spectrometry analysis as described under ‘Supplementary Experimental Procedures’.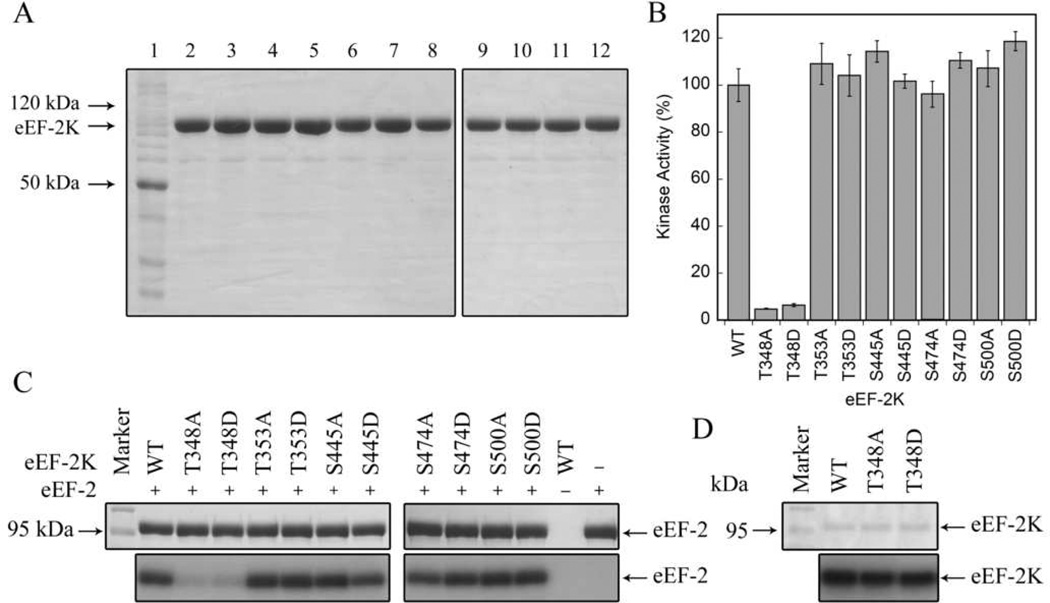


 - red) – suggested to be involved in the negative regulation of eEF-2K activity through an inhibitory phosphorylation (these sites include Ser-78, Ser-359, Ser-366 and Ser-396). Regulation through the mTOR pathway involves the phosphorylation of Ser-366 by p70 S6 kinase, and the phosphorylation of Ser-359 and Ser-78 by at least two additional unknown kinases (–24). It has been postulated that the Ser-78 phosphorylation acts to hinder the binding of CaM to eEF-2K (24). The cdc2-cyclin B complex has been shown to modulate eEF-2K activity via Ser-359 in a manner that is dependent on the cell cycle as well as amino acid availability, and is perhaps controlled by mTOR (25). Regulation through the MAPK cascade occurs via the phosphorylation of Ser-366 by p90RSK1 in an ERK-dependent fashion (22). In addition, the stress-activated protein kinases p38α and p38δ inhibit eEF2K via phosphorylation on Ser-396 (23). p38δ is also known to phosphorylate eEF-2K on Ser-359 (21); (
- red) – suggested to be involved in the negative regulation of eEF-2K activity through an inhibitory phosphorylation (these sites include Ser-78, Ser-359, Ser-366 and Ser-396). Regulation through the mTOR pathway involves the phosphorylation of Ser-366 by p70 S6 kinase, and the phosphorylation of Ser-359 and Ser-78 by at least two additional unknown kinases (–24). It has been postulated that the Ser-78 phosphorylation acts to hinder the binding of CaM to eEF-2K (24). The cdc2-cyclin B complex has been shown to modulate eEF-2K activity via Ser-359 in a manner that is dependent on the cell cycle as well as amino acid availability, and is perhaps controlled by mTOR (25). Regulation through the MAPK cascade occurs via the phosphorylation of Ser-366 by p90RSK1 in an ERK-dependent fashion (22). In addition, the stress-activated protein kinases p38α and p38δ inhibit eEF2K via phosphorylation on Ser-396 (23). p38δ is also known to phosphorylate eEF-2K on Ser-359 (21); ( - green) – suggested to be involved in the positive regulation of eEF-2K activity through an activating phosphorylation (these sites include Ser-398 and Ser-500). Phosphorylation of Ser-398 by the energy-supply regulator AMPK is known to activate eEF-2K (29). The cAMP-dependent PKA has also been shown to activate eEF-2K via a phosphorylation on Ser-500, and in the process imparts Ca2+-independent activity to the kinase (–28); (
- green) – suggested to be involved in the positive regulation of eEF-2K activity through an activating phosphorylation (these sites include Ser-398 and Ser-500). Phosphorylation of Ser-398 by the energy-supply regulator AMPK is known to activate eEF-2K (29). The cAMP-dependent PKA has also been shown to activate eEF-2K via a phosphorylation on Ser-500, and in the process imparts Ca2+-independent activity to the kinase (–28); ( - blue) – involved in autophosphorylation of eEF-2K (these sites include Thr-348, Thr-353, Ser-445, Ser-474 and Ser-500). Of the 5 autophosphorylation sites, only Thr-348 appears to be essential for activity against its substrate. Ser-500 is an autophosphorylation site and is also known to be phosphorylated by PKA, and could be the key residue responsible for autophosphorylation-induced Ca2+-independent (CaM-dependent – this work) activity (16, 17). The role of the phosphorylation at Ser-377 by MAPKAP-K2 has not yet been determined (23).
- blue) – involved in autophosphorylation of eEF-2K (these sites include Thr-348, Thr-353, Ser-445, Ser-474 and Ser-500). Of the 5 autophosphorylation sites, only Thr-348 appears to be essential for activity against its substrate. Ser-500 is an autophosphorylation site and is also known to be phosphorylated by PKA, and could be the key residue responsible for autophosphorylation-induced Ca2+-independent (CaM-dependent – this work) activity (16, 17). The role of the phosphorylation at Ser-377 by MAPKAP-K2 has not yet been determined (23).References
-
- Hershey JWB. Translational Control in Mammalian Cells. Annual Review of Biochemistry. 1991;60:717–755. - PubMed
-
- Morley SJ, Thomas G. Intracellular messengers and the control of protein synthesis. Pharmacology & Therapeutics. 1991;50:291–319. - PubMed
-
- Proud CG. Protein phosphorylation in translational control. Curr Top Cell Regul. 1992;32:243–369. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
