

Carbon monoxide: impact on remethylation/transsulfuration metabolism and its pathophysiologic implications
- PMID: 22331189
- PMCID: PMC3296020
- DOI: 10.1007/s00109-012-0875-2
Carbon monoxide: impact on remethylation/transsulfuration metabolism and its pathophysiologic implications
Abstract
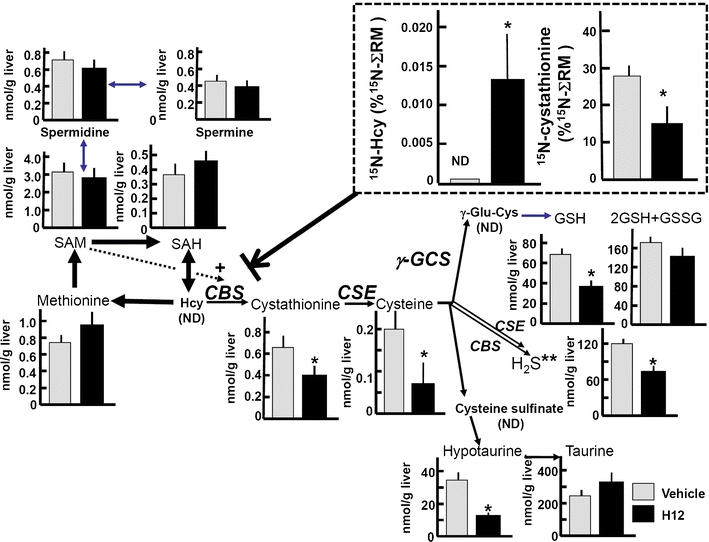
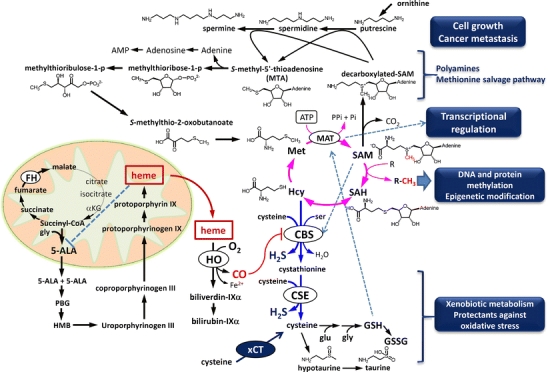
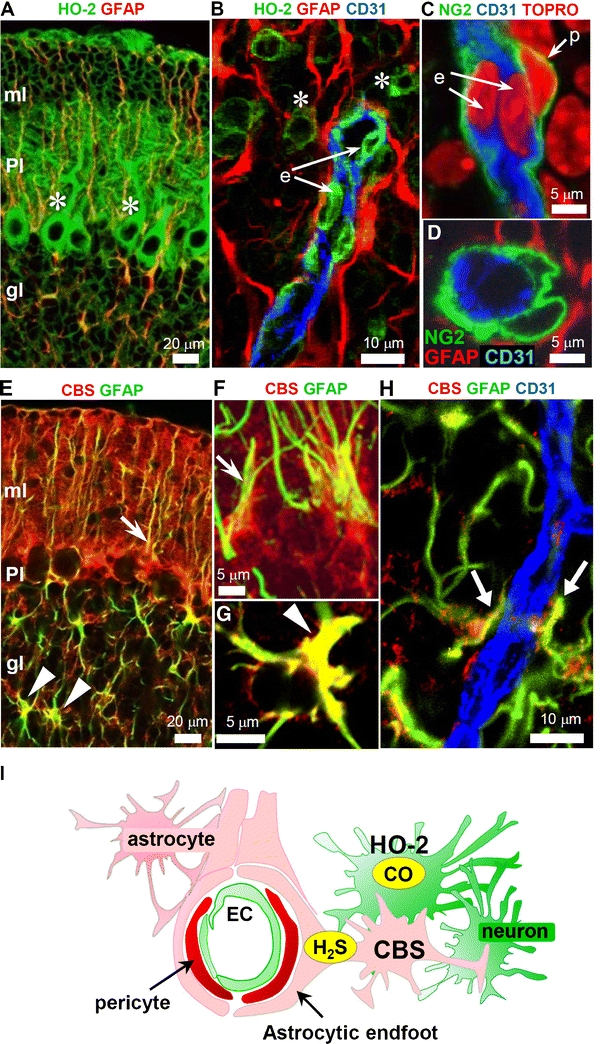
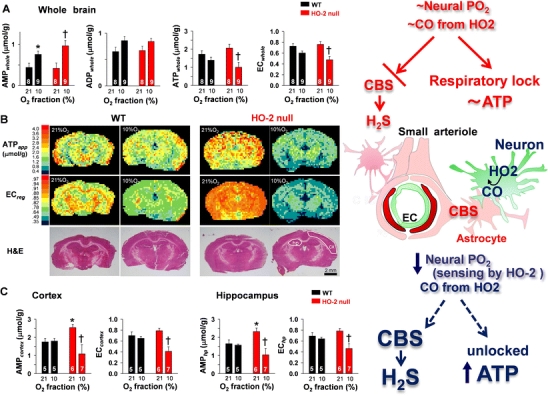
Carbon monoxide (CO) is a gaseous product generated by heme oxygenase (HO), which oxidatively degrades heme. While the stress-inducible HO-1 has well been recognized as an anti-oxidative defense mechanism under stress conditions, recent studies suggest that cancer cells utilize the reaction for their survival. HO-2, the constitutive isozyme, also plays protective roles as a tonic regulator for neurovascular function. Although protective roles of the enzyme reaction and CO have extensively been studied, little information is available on the molecular mechanisms by which the gas exerts its biological actions. Recent studies using metabolomics revealed that CO inhibits cystathionine β-synthase (CBS), which generates H(2)S, another gaseous mediator. The CO-dependent CBS inhibition may impact on the remethylation cycle and related metabolic pathways including the methionine salvage pathway and polyamine synthesis. This review focuses on the gas-responsive regulation of metabolic systems, particularly the remethylation and transsulfuration pathways, and their putative implications for cancer and ischemic diseases.
Figures

References
-
- Tenhunen R, Marver HS, Schmid R. Microsomal heme oxygenase. Characterization of the enzyme. J Biol Chem. 1969;244:6388–6394. - PubMed
-
- Yoshida T, Takahashi S, Kikuchi G. Partial purification and reconstitution of the heme oxygenase system from pig spleen microsomes. J Biochem. 1974;75:1187–1191. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
