

Size control of the Drosophila hematopoietic niche by bone morphogenetic protein signaling reveals parallels with mammals
- PMID: 22331866
- PMCID: PMC3295293
- DOI: 10.1073/pnas.1109407109
Size control of the Drosophila hematopoietic niche by bone morphogenetic protein signaling reveals parallels with mammals
Abstract
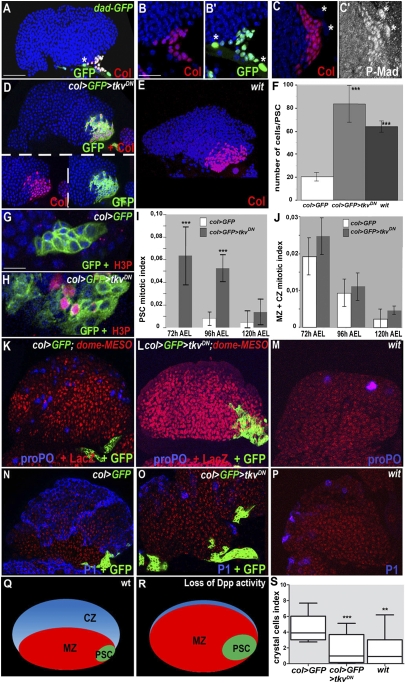
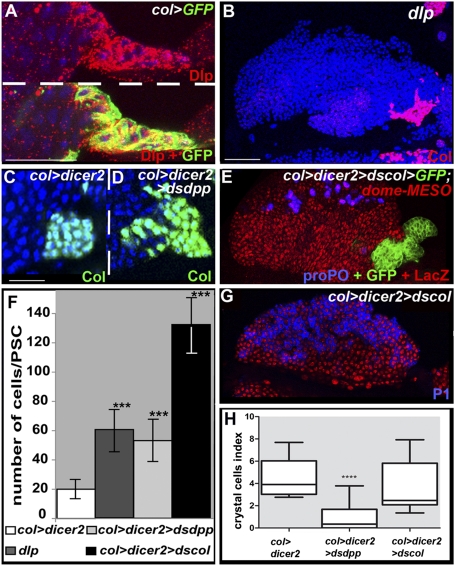
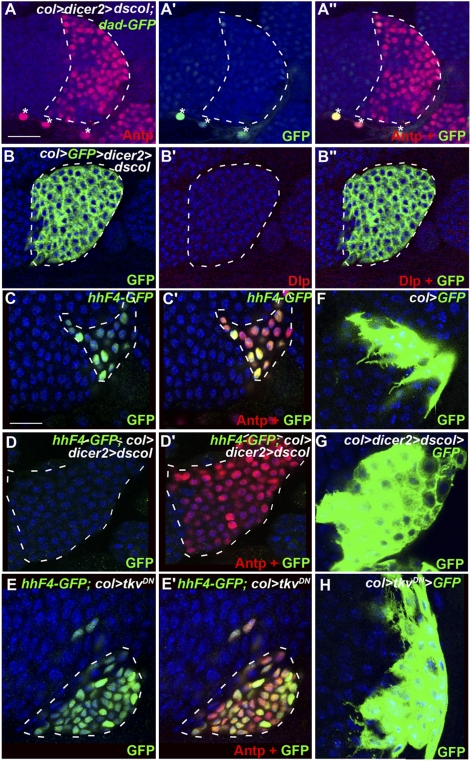
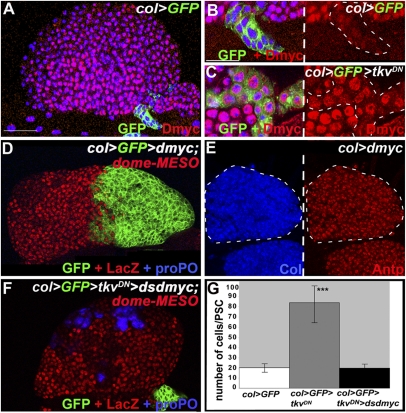
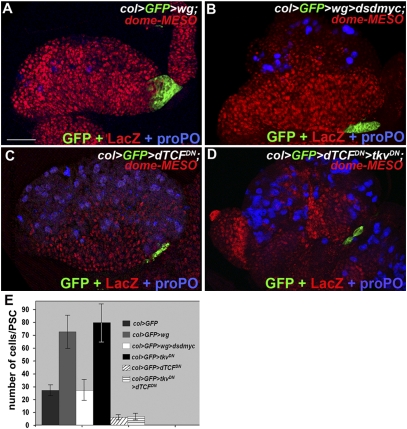
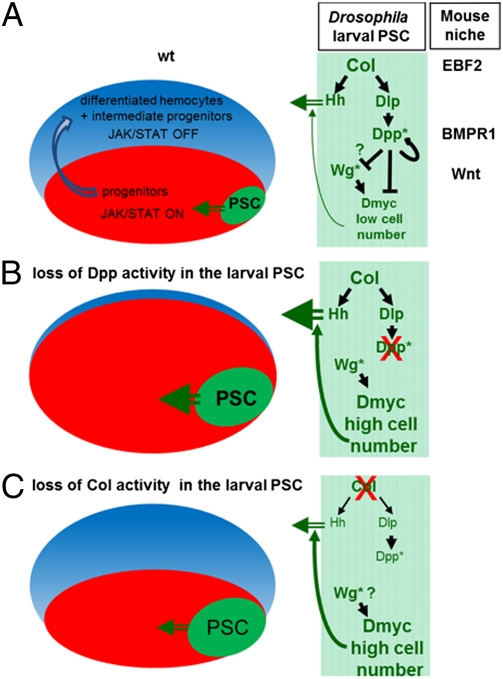
The Drosophila melanogaster larval hematopoietic organ, the lymph gland, is a model to study in vivo the function of the hematopoietic niche. A small cluster of cells in the lymph gland, the posterior signaling center (PSC), maintains the balance between hematopoietic progenitors (prohemocytes) and their differentiation into specialized blood cells (hemocytes). Here, we show that Decapentaplegic/bone morphogenetic protein (Dpp/BMP) signaling activity in PSC cells controls niche size. In the absence of BMP signaling, the number of PSC cells increases. Correlatively, no hemocytes differentiate. Controlling PSC size is, thus, essential for normal blood cell homeostasis. Activation of BMP signaling in the PSC requires expression of the Dally-like heparan-sulfate proteoglycan, under the control of the Collier/early B-cell factor (EBF) transcription factor. A Dpp > dpp autoregulatory loop maintains BMP signaling, which limits PSC cell proliferation by repressing the protooncogene dmyc. Dpp antagonizes activity of wingless (Wg)/Wnt signaling, which positively regulates the number of PSC cells via the control of Dmyc expression. Together, our data show that Collier controls hemocyte homeostasis via coordinate regulation of PSC cell number and PSC signaling to prohemocytes. In mouse, EBF2, BMP, and Wnt signaling in osteoblasts is required for the proper number of niche and hematopoietic stem cells. Our findings bring insights to niche size control and draw parallels between Drosophila and mammalian hematopoiesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Krzemien J, Crozatier M, Vincent A. Ontogeny of the Drosophila larval hematopoietic organ, hemocyte homeostasis and the dedicated cellular immune response to parasitism. Int J Dev Biol. 2010;54:1117–1125. - PubMed
-
- Lanot R, Zachary D, Holder F, Meister M. Postembryonic hematopoiesis in Drosophila. Dev Biol. 2001;230:243–257. - PubMed
-
- Jung SH, Evans CJ, Uemura C, Banerjee U. The Drosophila lymph gland as a developmental model of hematopoiesis. Development. 2005;132:2521–2533. - PubMed
-
- Krzemien J, Oyallon J, Crozatier M, Vincent A. Hematopoietic progenitors and hemocyte lineages in the Drosophila lymph gland. Dev Biol. 2010;346:310–319. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
