

Unified quantitative model of AMPA receptor trafficking at synapses
- PMID: 22331885
- PMCID: PMC3295262
- DOI: 10.1073/pnas.1109818109
Unified quantitative model of AMPA receptor trafficking at synapses
Abstract
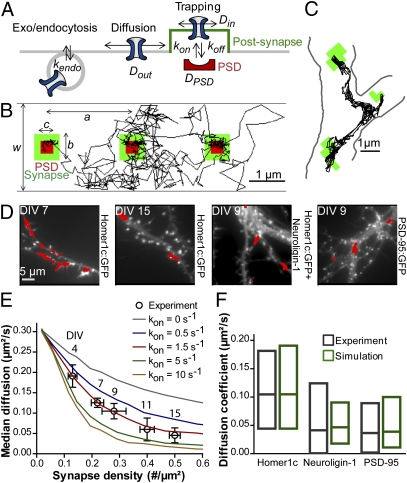
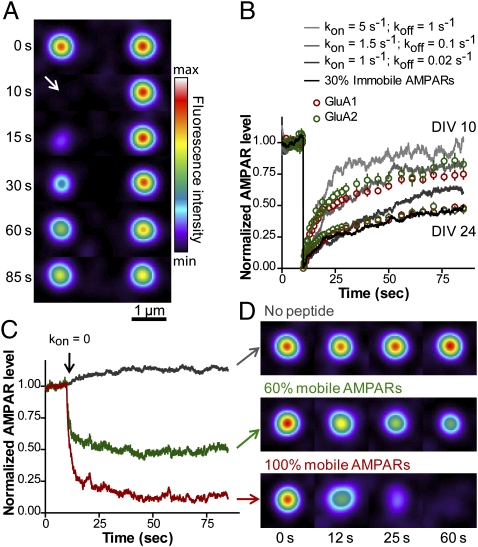
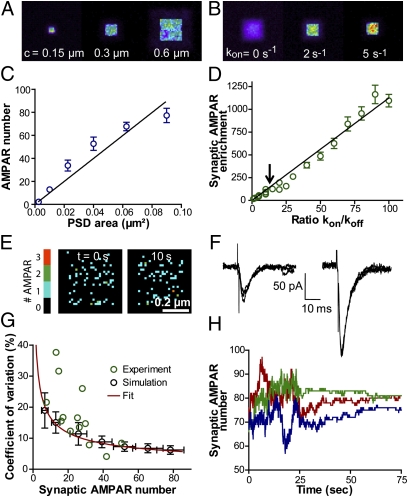
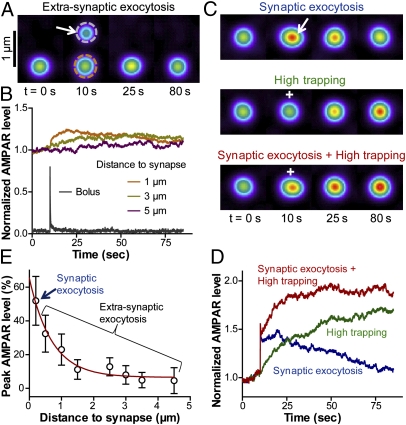
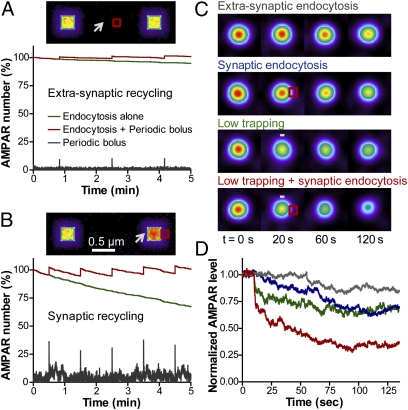
Trafficking of AMPA receptors (AMPARs) plays a key role in synaptic transmission. However, a general framework integrating the two major mechanisms regulating AMPAR delivery at postsynapses (i.e., surface diffusion and internal recycling) is lacking. To this aim, we built a model based on numerical trajectories of individual AMPARs, including free diffusion in the extrasynaptic space, confinement in the synapse, and trapping at the postsynaptic density (PSD) through reversible interactions with scaffold proteins. The AMPAR/scaffold kinetic rates were adjusted by comparing computer simulations to single-particle tracking and fluorescence recovery after photobleaching experiments in primary neurons, in different conditions of synapse density and maturation. The model predicts that the steady-state AMPAR number at synapses is bidirectionally controlled by AMPAR/scaffold binding affinity and PSD size. To reveal the impact of recycling processes in basal conditions and upon synaptic potentiation or depression, spatially and temporally defined exocytic and endocytic events were introduced. The model predicts that local recycling of AMPARs close to the PSD, coupled to short-range surface diffusion, provides rapid control of AMPAR number at synapses. In contrast, because of long-range diffusion limitations, extrasynaptic recycling is intrinsically slower and less synapse-specific. Thus, by discriminating the relative contributions of AMPAR diffusion, trapping, and recycling events on spatial and temporal bases, this model provides unique insights on the dynamic regulation of synaptic strength.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
