

Molecular mechanism of pharmacological activation of BK channels
- PMID: 22331907
- PMCID: PMC3295268
- DOI: 10.1073/pnas.1114321109
Molecular mechanism of pharmacological activation of BK channels
Abstract
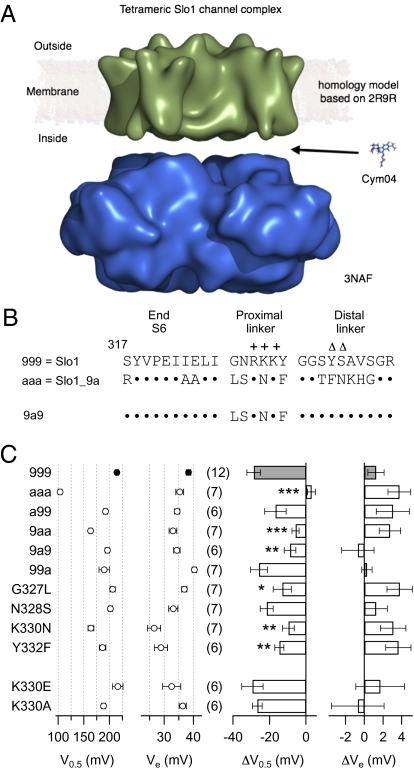
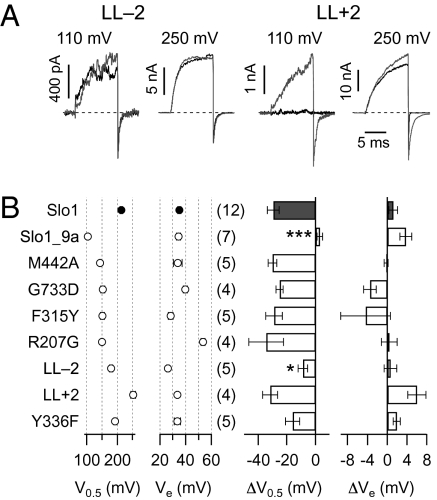
Large-conductance voltage- and Ca(2+)-activated K(+) (Slo1 BK) channels serve numerous cellular functions, and their dysregulation is implicated in various diseases. Drugs activating BK channels therefore bear substantial therapeutic potential, but their deployment has been hindered in part because the mode of action remains obscure. Here we provide mechanistic insight into how the dehydroabietic acid derivative Cym04 activates BK channels. As a representative of NS1619-like BK openers, Cym04 reversibly left-shifts the half-activation voltage of Slo1 BK channels. Using an established allosteric BK gating model, the Cym04 effect can be simulated by a shift of the voltage sensor and the ion conduction gate equilibria toward the activated and open state, respectively. BK activation by Cym04 occurs in a splice variant-specific manner; it does not occur in such Slo1 BK channels using an alternative neuronal exon 9, which codes for the linker connecting the transmembrane segment S6 and the cytosolic RCK1 domain--the S6/RCK linker. In addition, Cym04 does not affect Slo1 BK channels with a two-residue deletion within this linker. Mutagenesis and model-based gating analysis revealed that BK openers, such as Cym04 and NS1619 but not mallotoxin, activate BK channels by functionally interacting with the S6/RCK linker, mimicking site-specific shortening of this purported passive spring, which transmits force from the cytosolic gating ring structure to open the channel's gate.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
Modulation of BK channel gating by the ß2 subunit involves both membrane-spanning and cytoplasmic domains of Slo1.J Neurosci. 2010 Dec 1;30(48):16170-9. doi: 10.1523/JNEUROSCI.2323-10.2010. J Neurosci. 2010. PMID: 21123563 Free PMC article.
-
A point mutation in the human Slo1 channel that impairs its sensitivity to omega-3 docosahexaenoic acid.J Gen Physiol. 2013 Nov;142(5):507-22. doi: 10.1085/jgp.201311061. Epub 2013 Oct 14. J Gen Physiol. 2013. PMID: 24127525 Free PMC article.
-
Properties of Slo1 K+ channels with and without the gating ring.Proc Natl Acad Sci U S A. 2013 Oct 8;110(41):16657-62. doi: 10.1073/pnas.1313433110. Epub 2013 Sep 25. Proc Natl Acad Sci U S A. 2013. PMID: 24067659 Free PMC article.
-
Transduction of voltage and Ca2+ signals by Slo1 BK channels.Physiology (Bethesda). 2013 May;28(3):172-89. doi: 10.1152/physiol.00055.2012. Physiology (Bethesda). 2013. PMID: 23636263 Free PMC article. Review.
-
Modulation of BK Channels by Small Endogenous Molecules and Pharmaceutical Channel Openers.Int Rev Neurobiol. 2016;128:193-237. doi: 10.1016/bs.irn.2016.03.020. Epub 2016 May 4. Int Rev Neurobiol. 2016. PMID: 27238265 Review.
Cited by
-
KCNMA1-linked channelopathy.J Gen Physiol. 2019 Oct 7;151(10):1173-1189. doi: 10.1085/jgp.201912457. Epub 2019 Aug 19. J Gen Physiol. 2019. PMID: 31427379 Free PMC article. Review.
-
The smooth muscle-type β1 subunit potentiates activation by DiBAC4(3) in recombinant BK channels.Channels (Austin). 2014;8(1):95-102. doi: 10.4161/chan.27212. Epub 2013 Dec 3. Channels (Austin). 2014. PMID: 24299688 Free PMC article.
-
Atomistic Simulation of Voltage Activation of a Truncated BK Channel.bioRxiv [Preprint]. 2025 Jun 14:2025.01.08.631907. doi: 10.1101/2025.01.08.631907. bioRxiv. 2025. PMID: 40661448 Free PMC article. Preprint.
-
Single-channel biophysical and pharmacological characterizations of native human large-conductance calcium-activated potassium channels in freshly isolated detrusor smooth muscle cells.Pflugers Arch. 2013 Jul;465(7):965-75. doi: 10.1007/s00424-012-1214-8. Epub 2013 Jan 24. Pflugers Arch. 2013. PMID: 23344746 Free PMC article.
-
Pro-inflammatory Cytokines Drive Deregulation of Potassium Channel Expression in Primary Synovial Fibroblasts.Front Physiol. 2020 Mar 24;11:226. doi: 10.3389/fphys.2020.00226. eCollection 2020. Front Physiol. 2020. PMID: 32265733 Free PMC article.
References
-
- Salkoff L, Butler A, Ferreira G, Santi C, Wei A. High-conductance potassium channels of the SLO family. Nat Rev Neurosci. 2006;7:921–931. - PubMed
-
- Jaggar JH, Porter VA, Lederer WJ, Nelson MT. Calcium sparks in smooth muscle. Am J Physiol Cell Physiol. 2000;278:C235–C256. - PubMed
-
- Orio P, Rojas P, Ferreira G, Latorre R. New disguises for an old channel: MaxiK channel β-subunits. News Physiol Sci. 2002;17:156–161. - PubMed
-
- Fodor AA, Aldrich RW. Convergent evolution of alternative splices at domain boundaries of the BK channel. Annu Rev Physiol. 2009;71:19–36. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
