

Computational analysis of pathogen-borne metallo β-lactamases reveals discriminating structural features between B1 types
- PMID: 22333139
- PMCID: PMC3293060
- DOI: 10.1186/1756-0500-5-96
Computational analysis of pathogen-borne metallo β-lactamases reveals discriminating structural features between B1 types
Abstract
Background: Genes conferring antibiotic resistance to groups of bacterial pathogens are cause for considerable concern, as many once-reliable antibiotics continue to see a reduction in efficacy. The recent discovery of the metallo β-lactamase blaNDM-1 gene, which appears to grant antibiotic resistance to a variety of Enterobacteriaceae via a mobile plasmid, is one example of this distressing trend. The following work describes a computational analysis of pathogen-borne MBLs that focuses on the structural aspects of characterized proteins.
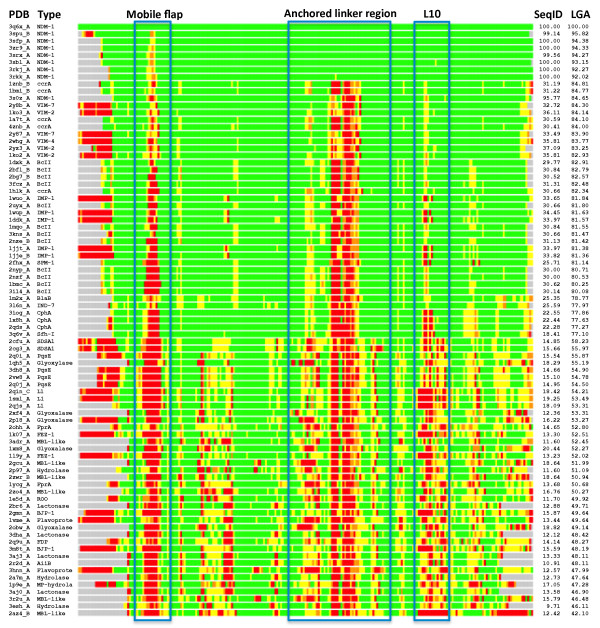
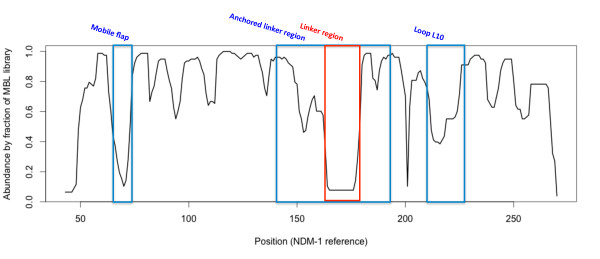
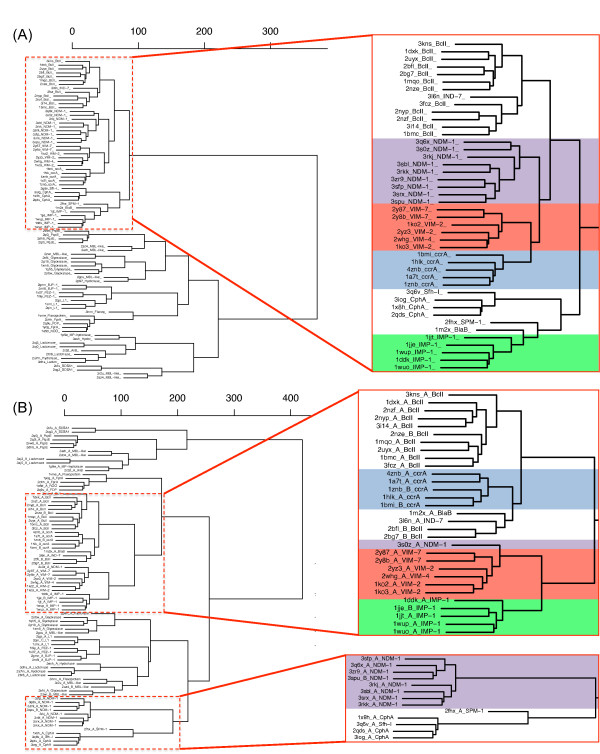
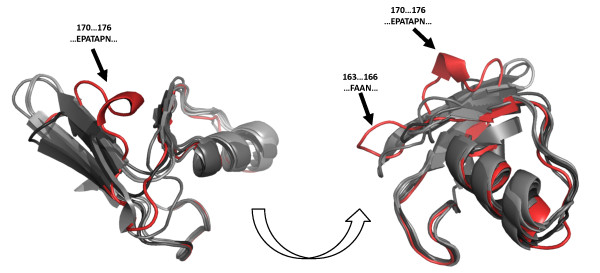
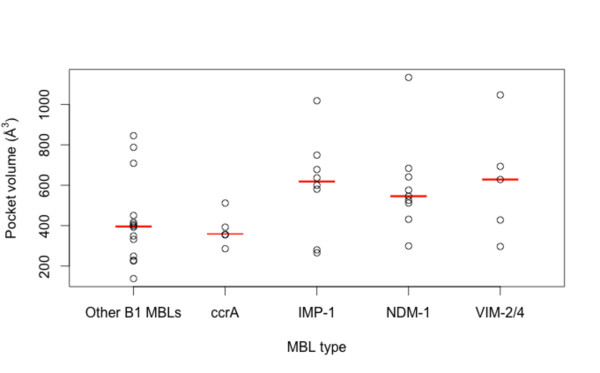
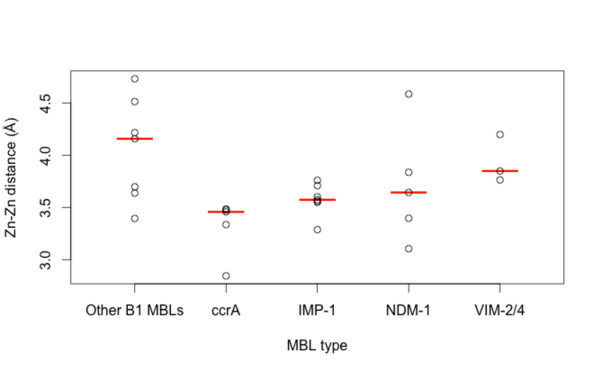
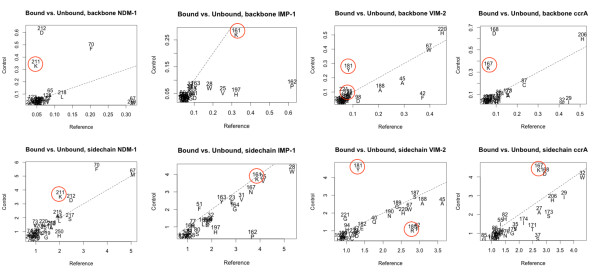
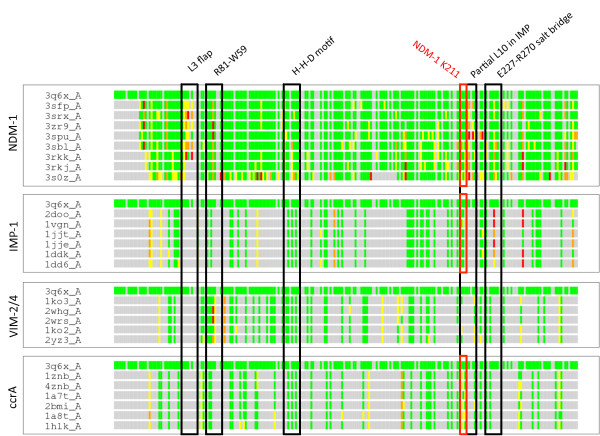
Results: Using both sequence and structural analyses, we examine residues and structural features specific to various pathogen-borne MBL types. This analysis identifies a linker region within MBL-like folds that may act as a discriminating structural feature between these proteins, and specifically resistance-associated acquirable MBLs. Recently released crystal structures of the newly emerged NDM-1 protein were aligned against related MBL structures using a variety of global and local structural alignment methods, and the overall fold conformation is examined for structural conservation. Conservation appears to be present in most areas of the protein, yet is strikingly absent within a linker region, making NDM-1 unique with respect to a linker-based classification scheme. Variability analysis of the NDM-1 crystal structure highlights unique residues in key regions as well as identifying several characteristics shared with other transferable MBLs.
Conclusions: A discriminating linker region identified in MBL proteins is highlighted and examined in the context of NDM-1 and primarily three other MBL types: IMP-1, VIM-2 and ccrA. The presence of an unusual linker region variant and uncommon amino acid composition at specific structurally important sites may help to explain the unusually broad kinetic profile of NDM-1 and may aid in directing research attention to areas of this protein, and possibly other MBLs, that may be targeted for inactivation or attenuation of enzymatic activity.
Figures

Similar articles
-
Sulfamoyl Heteroarylcarboxylic Acids as Promising Metallo-β-Lactamase Inhibitors for Controlling Bacterial Carbapenem Resistance.mBio. 2020 Mar 17;11(2):e03144-19. doi: 10.1128/mBio.03144-19. mBio. 2020. PMID: 32184250 Free PMC article.
-
Multiyear, Multinational Survey of the Incidence and Global Distribution of Metallo-β-Lactamase-Producing Enterobacteriaceae and Pseudomonas aeruginosa.Antimicrob Agents Chemother. 2015 Dec 7;60(2):1067-78. doi: 10.1128/AAC.02379-15. Print 2016 Feb. Antimicrob Agents Chemother. 2015. PMID: 26643349 Free PMC article.
-
Probing the Interaction of Aspergillomarasmine A with Metallo-β-lactamases NDM-1, VIM-2, and IMP-7.ACS Infect Dis. 2018 Feb 9;4(2):135-145. doi: 10.1021/acsinfecdis.7b00106. Epub 2017 Nov 9. ACS Infect Dis. 2018. PMID: 29091730 Free PMC article.
-
Epidemiology and Characteristics of Metallo-β-Lactamase-Producing Pseudomonas aeruginosa.Infect Chemother. 2015 Jun;47(2):81-97. doi: 10.3947/ic.2015.47.2.81. Epub 2015 Jun 30. Infect Chemother. 2015. PMID: 26157586 Free PMC article. Review.
-
Acquired metallo-β-lactamases and their genetic association with class 1 integrons and ISCR elements in Gram-negative bacteria.Future Microbiol. 2015;10(5):873-87. doi: 10.2217/fmb.15.18. Future Microbiol. 2015. PMID: 26000655 Review.
Cited by
-
The mechanism of NDM-1-catalyzed carbapenem hydrolysis is distinct from that of penicillin or cephalosporin hydrolysis.Nat Commun. 2017 Dec 21;8(1):2242. doi: 10.1038/s41467-017-02339-w. Nat Commun. 2017. PMID: 29269938 Free PMC article.
-
A case study comparing quantitative stability-flexibility relationships across five metallo-β-lactamases highlighting differences within NDM-1.Methods Mol Biol. 2014;1084:227-38. doi: 10.1007/978-1-62703-658-0_12. Methods Mol Biol. 2014. PMID: 24061924 Free PMC article.
-
Molecular diversity of extended-spectrum β-lactamases and carbapenemases, and antimicrobial resistance.J Intensive Care. 2020 Jan 28;8:13. doi: 10.1186/s40560-020-0429-6. eCollection 2020. J Intensive Care. 2020. PMID: 32015881 Free PMC article. Review.
-
Structural Basis of Metallo-β-Lactamase Inhibition by Captopril Stereoisomers.Antimicrob Agents Chemother. 2015 Oct 19;60(1):142-50. doi: 10.1128/AAC.01335-15. Print 2016 Jan. Antimicrob Agents Chemother. 2015. PMID: 26482303 Free PMC article.
-
The interaction of the azetidine thiazole side chain with the active site loop (ASL) 3 drives the evolution of IMP metallo-β-lactamase against tebipenem.Antimicrob Agents Chemother. 2024 Aug 7;68(8):e0068724. doi: 10.1128/aac.00687-24. Epub 2024 Jul 18. Antimicrob Agents Chemother. 2024. PMID: 39023262 Free PMC article.
References
-
- Lee K, Yum JH, Yong D, Lee HM, Kim HD, Docquier J-D, Rossolini GM, Chong Y. Novel acquired metallo-beta-lactamase gene, bla(sim-1), in a class 1 integron from acinetobacter baumannii clinical isolates from Korea. Antimicrob Agents Chemother. 2005;49(11):4485–4491. doi: 10.1128/AAC.49.11.4485-4491.2005. - DOI - PMC - PubMed
-
- Yong D, Toleman MA, Giske CG, Cho HS, Sundman K, Lee K, Walsh TR. Characterization of a new metallo-beta-lactamase gene, blaNDM-1, and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob Agents Chemother. 2009;53(12):5046–5054. doi: 10.1128/AAC.00774-09. - DOI - PMC - PubMed
-
- Grundmann H, Livermore DM, Giske CG, Canton R, Rossolini GM, Campos J, Vatopoulos A, Gniadkowski M, Toth A, Pfeifer Y, Jarlier V, Carmeli Y. CNSE Working Group. Carbapenem-non-susceptible enterobacteriaceae in Europe: conclusions from a meeting of national experts. Euro Surveill. 2010;15(46):1–13. - PubMed
-
- Kumarasamy KK, Toleman MA, Walsh TR, Bagaria J, Butt F, Balakrishnan R, Chaudhary U, Doumith M, Giske CG, Irfan S, Krishnan P, Kumar AV, Maharjan S, Mushtaq S, Noorie T, Paterson DL, Pearson A, Perry C, Pike R, Rao B, Ray U, Sarma JB, Sharma M, Sheridan E, Thirunarayan MA, Turton J, Upadhyay S, Warner M, Welfare W, Livermore DM, Woodford N. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: a molecular, biological, and epidemiological study. Lancet Infect Dis. 2010;10(9):597–602. doi: 10.1016/S1473-3099(10)70143-2. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
