

Genetic strain differences in learned fear inhibition associated with variation in neuroendocrine, autonomic, and amygdala dendritic phenotypes
- PMID: 22334122
- PMCID: PMC3327858
- DOI: 10.1038/npp.2011.340
Genetic strain differences in learned fear inhibition associated with variation in neuroendocrine, autonomic, and amygdala dendritic phenotypes
Abstract
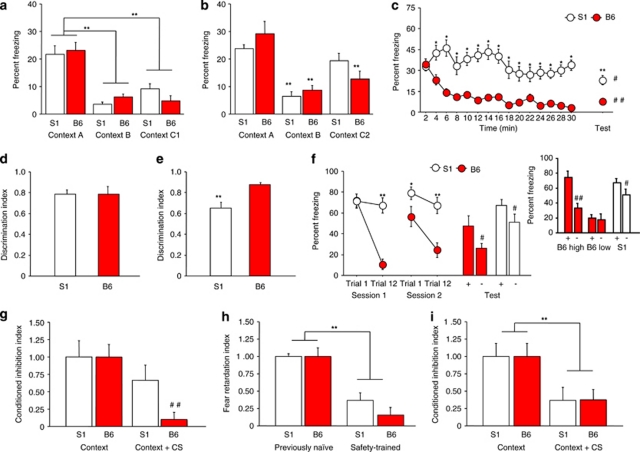
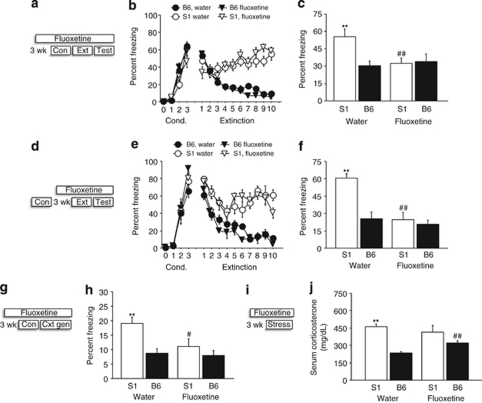
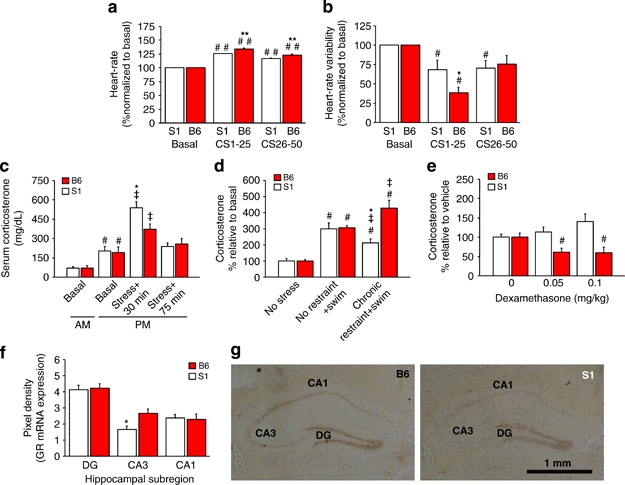
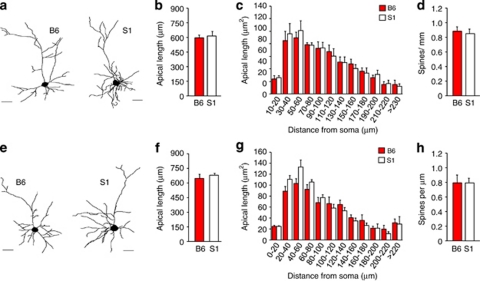
Mood and anxiety disorders develop in some but not all individuals following exposure to stress and psychological trauma. However, the factors underlying individual differences in risk and resilience for these disorders, including genetic variation, remain to be determined. Isogenic inbred mouse strains provide a valuable approach to elucidating these factors. Here, we performed a comprehensive examination of the extinction-impaired 129S1/SvImJ (S1) inbred mouse strain for multiple behavioral, autonomic, neuroendocrine, and corticolimbic neuronal morphology phenotypes. We found that S1 exhibited fear overgeneralization to ambiguous contexts and cues, impaired context extinction and impaired safety learning, relative to the (good-extinguishing) C57BL/6J (B6) strain. Fear overgeneralization and impaired extinction was rescued by treatment with the front-line anxiety medication fluoxetine. Telemetric measurement of electrocardiogram signals demonstrated autonomic disturbances in S1 including poor recovery of fear-induced suppression of heart rate variability. S1 with a history of chronic restraint stress displayed an attenuated corticosterone (CORT) response to a novel, swim stressor. Conversely, previously stress-naive S1 showed exaggerated CORT responses to acute restraint stress or extinction training, insensitivity to dexamethasone challenge, and reduced hippocampal CA3 glucocorticoid receptor mRNA, suggesting downregulation of negative feedback control of the hypothalamic-pituitary-adrenal axis. Analysis of neuronal morphology in key neural nodes within the fear and extinction circuit revealed enlarged dendritic arbors in basolateral amygdala neurons in S1, but normal infralimbic cortex and prelimbic cortex dendritic arborization. Collectively, these data provide convergent support for the utility of the S1 strain as a tractable model for elucidating the neural, molecular and genetic basis of persistent, excessive fear.
Figures

Similar articles
-
Prefrontal single-unit firing associated with deficient extinction in mice.Neurobiol Learn Mem. 2014 Sep;113:69-81. doi: 10.1016/j.nlm.2013.11.002. Epub 2013 Nov 11. Neurobiol Learn Mem. 2014. PMID: 24231425 Free PMC article.
-
Increased amygdalar metabotropic glutamate receptor 7 mRNA in a genetic mouse model of impaired fear extinction.Psychopharmacology (Berl). 2019 Jan;236(1):265-272. doi: 10.1007/s00213-018-5031-4. Epub 2018 Sep 13. Psychopharmacology (Berl). 2019. PMID: 30215216 Free PMC article.
-
Impaired fear extinction learning and cortico-amygdala circuit abnormalities in a common genetic mouse strain.J Neurosci. 2008 Aug 6;28(32):8074-85. doi: 10.1523/JNEUROSCI.4904-07.2008. J Neurosci. 2008. PMID: 18685032 Free PMC article.
-
Sleep-wake and arousal dysfunctions in post-traumatic stress disorder: Role of orexin systems.Brain Res Bull. 2022 Aug;186:106-122. doi: 10.1016/j.brainresbull.2022.05.006. Epub 2022 May 23. Brain Res Bull. 2022. PMID: 35618150 Review.
-
Rodent models of impaired fear extinction.Psychopharmacology (Berl). 2019 Jan;236(1):21-32. doi: 10.1007/s00213-018-5054-x. Epub 2018 Oct 31. Psychopharmacology (Berl). 2019. PMID: 30377749 Free PMC article. Review.
Cited by
-
Assessing negative affect in mice during abstinence from alcohol drinking: Limitations and future challenges.Alcohol. 2022 May;100:41-56. doi: 10.1016/j.alcohol.2022.02.001. Epub 2022 Feb 15. Alcohol. 2022. PMID: 35181404 Free PMC article. Review.
-
Animal Models of PTSD: The Socially Isolated Mouse and the Biomarker Role of Allopregnanolone.Front Behav Neurosci. 2019 Jun 11;13:114. doi: 10.3389/fnbeh.2019.00114. eCollection 2019. Front Behav Neurosci. 2019. PMID: 31244621 Free PMC article. Review.
-
Chronic inhibition of GABA synthesis in the infralimbic cortex facilitates conditioned safety memory and reduces contextual fear.Transl Psychiatry. 2020 Apr 24;10(1):120. doi: 10.1038/s41398-020-0788-8. Transl Psychiatry. 2020. PMID: 32332716 Free PMC article.
-
Sex differences in avoidance behavior and cued threat memory dynamics in mice: Interactions between estrous cycle and genetic background.Horm Behav. 2023 Nov;156:105439. doi: 10.1016/j.yhbeh.2023.105439. Epub 2023 Oct 7. Horm Behav. 2023. PMID: 37813043 Free PMC article.
-
Intranasal oxytocin compensates for estrus cycle-specific reduction of conditioned safety memory in rats: Implications for psychiatric disorders.Neurobiol Stress. 2021 Mar 10;14:100313. doi: 10.1016/j.ynstr.2021.100313. eCollection 2021 May. Neurobiol Stress. 2021. PMID: 33778132 Free PMC article.
References
-
- Arditi-Babchuk H, Feldman R, Gilboa-Schechtman E. Parasympathetic reactivity to recalled traumatic and pleasant events in trauma-exposed individuals. J Trauma Stress. 2009;22:254–257. - PubMed
-
- Barrick CJ, Rojas M, Schoonhoven R, Smyth SS, Threadgill DW. Cardiac response to pressure overload in 129S1/SvImJ and C57BL/6J mice: temporal- and background-dependent development of concentric left ventricular hypertrophy. Am J Physiol Heart Circ Physiol. 2007;292:H2119–H2130. - PubMed
-
- Blanchard RJ, Blanchard DC. Crouching as an index of fear. J Comp Physiol Psychol. 1969;67:370–375. - PubMed
-
- Bonne O, Grillon C, Vythilingam M, Neumeister A, Charney DS. Adaptive and maladaptive psychobiological responses to severe psychological stress: implications for the discovery of novel pharmacotherapy. Neurosci Biobehav Rev. 2004;28:65–94. - PubMed
-
- Boyce-Rustay JM, Cameron HA, Holmes A. Chronic swim stress alters sensitivity to acute behavioral effects of ethanol in mice. Physiol Behav. 2007;91:77–86. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
