

Monoubiquitinated histone H2A destabilizes photolesion-containing nucleosomes with concomitant release of UV-damaged DNA-binding protein E3 ligase
- PMID: 22334663
- PMCID: PMC3320950
- DOI: 10.1074/jbc.M111.307058
Monoubiquitinated histone H2A destabilizes photolesion-containing nucleosomes with concomitant release of UV-damaged DNA-binding protein E3 ligase
Abstract
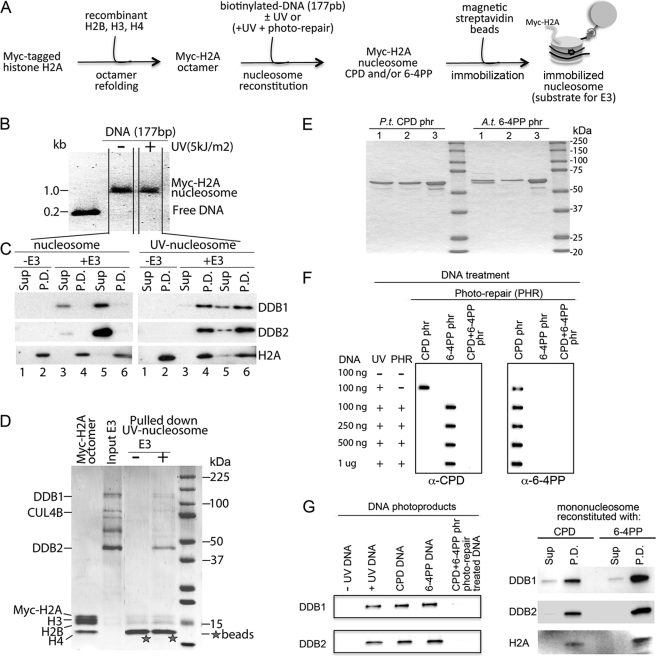
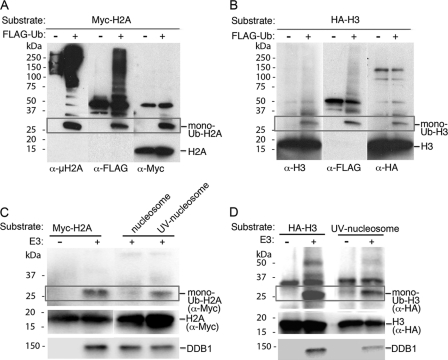
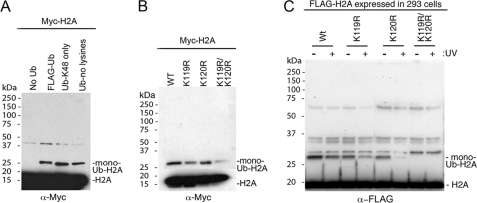
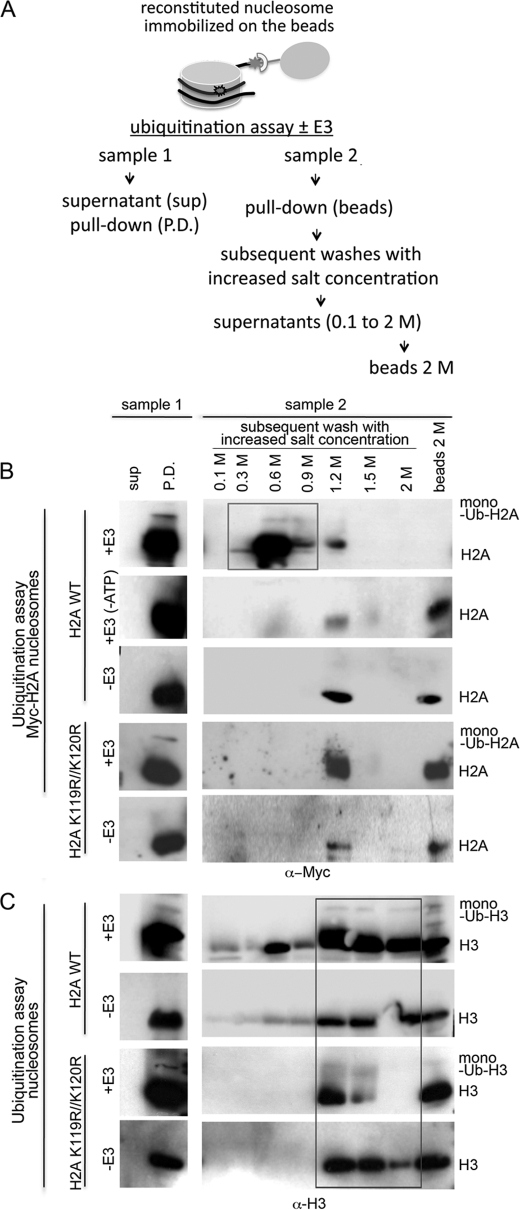
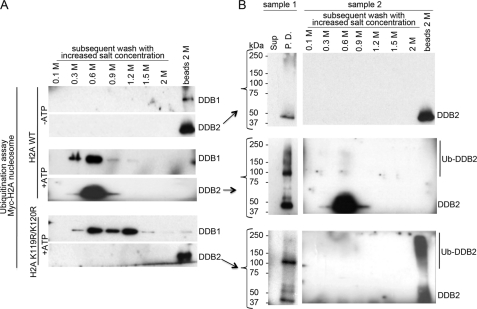
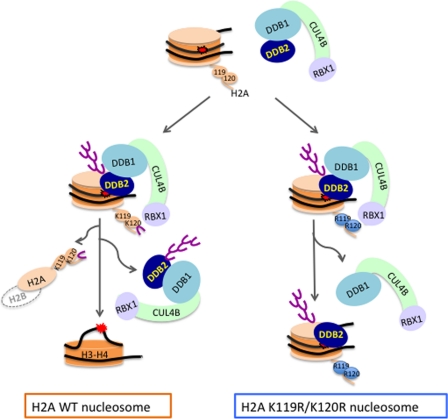
How the nucleotide excision repair (NER) machinery gains access to damaged chromatinized DNA templates and how the chromatin structure is modified to promote efficient repair of the non-transcribed genome remain poorly understood. The UV-damaged DNA-binding protein complex (UV-DDB, consisting of DDB1 and DDB2, the latter of which is mutated in xeroderma pigmentosum group E patients, is a substrate-recruiting module of the cullin 4B-based E3 ligase complex, DDB1-CUL4B(DDB2). We previously reported that the deficiency of UV-DDB E3 ligases in ubiquitinating histone H2A at UV-damaged DNA sites in the xeroderma pigmentosum group E cells contributes to the faulty NER in these skin cancer-prone patients. Here, we reveal the mechanism by which monoubiquitination of specific H2A lysine residues alters nucleosomal dynamics and subsequently initiates NER. We show that DDB1-CUL4B(DDB2) E3 ligase specifically binds to mononucleosomes assembled with human recombinant histone octamers and nucleosome-positioning DNA containing cyclobutane pyrimidine dimers or 6-4 photoproducts photolesions. We demonstrate functionally that ubiquitination of H2A Lys-119/Lys-120 is necessary for destabilization of nucleosomes and concomitant release of DDB1-CUL4B(DDB2) from photolesion-containing DNA. Nucleosomes in which these lysines are replaced with arginines are resistant to such structural changes, and arginine mutants prevent the eviction of H2A and dissociation of polyubiquitinated DDB2 from UV-damaged nucleosomes. The partial eviction of H3 from the nucleosomes is dependent on ubiquitinated H2A Lys-119/Lys-120. Our results provide mechanistic insight into how post-translational modification of H2A at the site of a photolesion initiates the repair process and directly affects the stability of the human genome.
Figures
Similar articles
-
Expanding molecular roles of UV-DDB: Shining light on genome stability and cancer.DNA Repair (Amst). 2020 Oct;94:102860. doi: 10.1016/j.dnarep.2020.102860. Epub 2020 Apr 27. DNA Repair (Amst). 2020. PMID: 32739133 Free PMC article. Review.
-
The cullin 4B-based UV-damaged DNA-binding protein ligase binds to UV-damaged chromatin and ubiquitinates histone H2A.Cancer Res. 2008 Jul 1;68(13):5014-22. doi: 10.1158/0008-5472.CAN-07-6162. Cancer Res. 2008. PMID: 18593899
-
The DDB1-CUL4ADDB2 ubiquitin ligase is deficient in xeroderma pigmentosum group E and targets histone H2A at UV-damaged DNA sites.Proc Natl Acad Sci U S A. 2006 Feb 21;103(8):2588-93. doi: 10.1073/pnas.0511160103. Epub 2006 Feb 10. Proc Natl Acad Sci U S A. 2006. PMID: 16473935 Free PMC article.
-
DNA damage binding protein component DDB1 participates in nucleotide excision repair through DDB2 DNA-binding and cullin 4A ubiquitin ligase activity.Cancer Res. 2006 Sep 1;66(17):8590-7. doi: 10.1158/0008-5472.CAN-06-1115. Cancer Res. 2006. PMID: 16951172
-
Open, repair and close again: chromatin dynamics and the response to UV-induced DNA damage.DNA Repair (Amst). 2011 Feb 7;10(2):119-25. doi: 10.1016/j.dnarep.2010.10.010. Epub 2010 Dec 3. DNA Repair (Amst). 2011. PMID: 21130713 Review.
Cited by
-
Role of poly(ADP-ribose) polymerase-1 in the removal of UV-induced DNA lesions by nucleotide excision repair.Proc Natl Acad Sci U S A. 2013 Jan 29;110(5):1658-63. doi: 10.1073/pnas.1209507110. Epub 2013 Jan 14. Proc Natl Acad Sci U S A. 2013. PMID: 23319653 Free PMC article.
-
Real-Time Tracking of Parental Histones Reveals Their Contribution to Chromatin Integrity Following DNA Damage.Mol Cell. 2016 Oct 6;64(1):65-78. doi: 10.1016/j.molcel.2016.08.019. Epub 2016 Sep 15. Mol Cell. 2016. PMID: 27642047 Free PMC article.
-
Histone H2A variants in nucleosomes and chromatin: more or less stable?Nucleic Acids Res. 2012 Nov;40(21):10719-41. doi: 10.1093/nar/gks865. Epub 2012 Sep 21. Nucleic Acids Res. 2012. PMID: 23002134 Free PMC article. Review.
-
Expanding molecular roles of UV-DDB: Shining light on genome stability and cancer.DNA Repair (Amst). 2020 Oct;94:102860. doi: 10.1016/j.dnarep.2020.102860. Epub 2020 Apr 27. DNA Repair (Amst). 2020. PMID: 32739133 Free PMC article. Review.
-
Global and transcription-coupled repair of 8-oxoG is initiated by nucleotide excision repair proteins.Nat Commun. 2022 Feb 21;13(1):974. doi: 10.1038/s41467-022-28642-9. Nat Commun. 2022. PMID: 35190564 Free PMC article.
References
-
- Friedberg E. C., Walker G. C., Siede W., Wood R. D., Schultz R. A., Ellenberger T. (2005) DNA Repair and Mutagenesis, 2nd Edition, American Society for Microbiology, Washington, D. C
-
- Gillet L. C., Schärer O. D. (2006) Molecular mechanisms of mammalian global genome nucleotide excision repair. Chem. Rev. 106, 253–276 - PubMed
-
- Naegeli H., Sugasawa K. (2011) The xeroderma pigmentosum pathway. Decision tree analysis of DNA quality. DNA Repair 10, 673–683 - PubMed
-
- Reed S. H. (2011) Nucleotide excision repair in chromatin. Damage removal at the drop of a HAT. DNA Repair 10, 734–742 - PubMed
-
- Luger K., Mäder A. W., Richmond R. K., Sargent D. F., Richmond T. J. (1997) Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 389, 251–260 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
