

Engineering mammalian mucin-type O-glycosylation in plants
- PMID: 22334671
- PMCID: PMC3320939
- DOI: 10.1074/jbc.M111.312918
Engineering mammalian mucin-type O-glycosylation in plants
Abstract
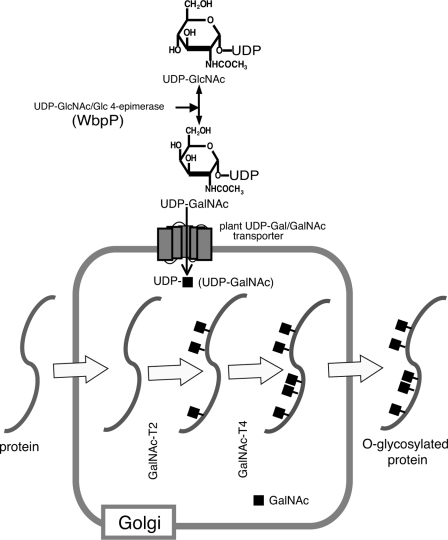
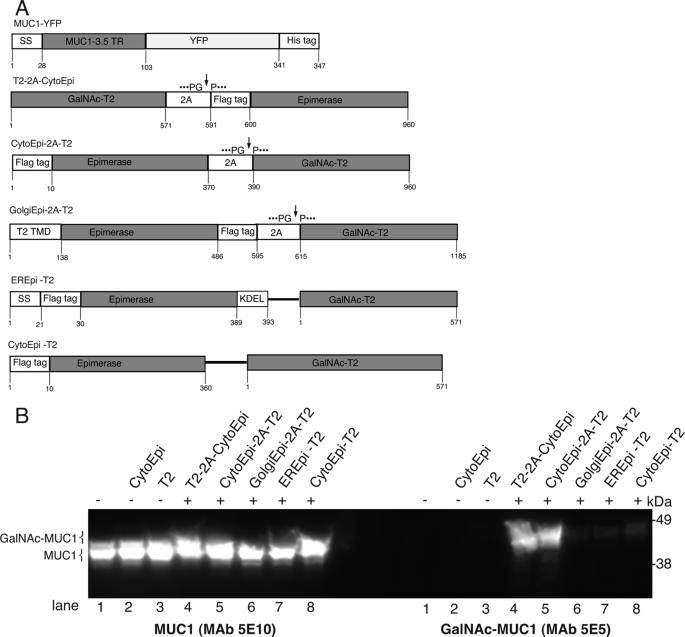
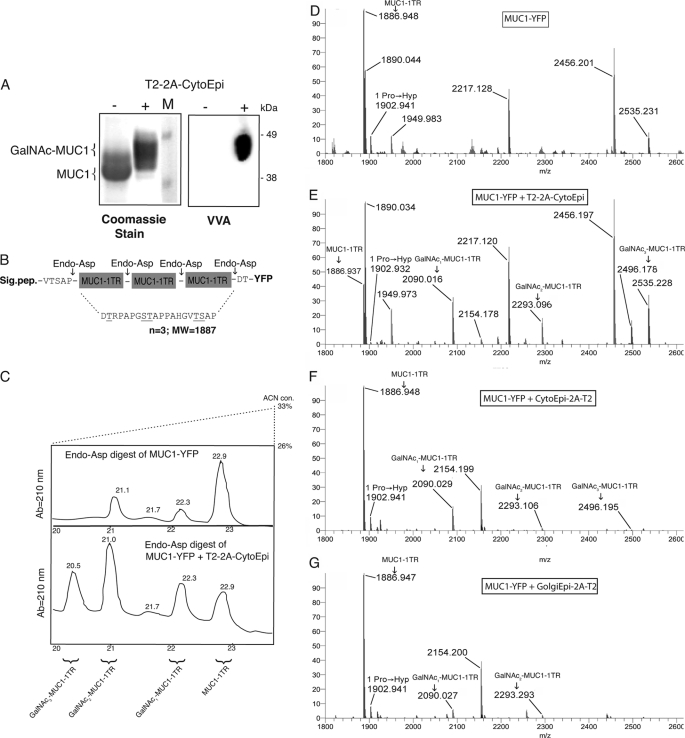
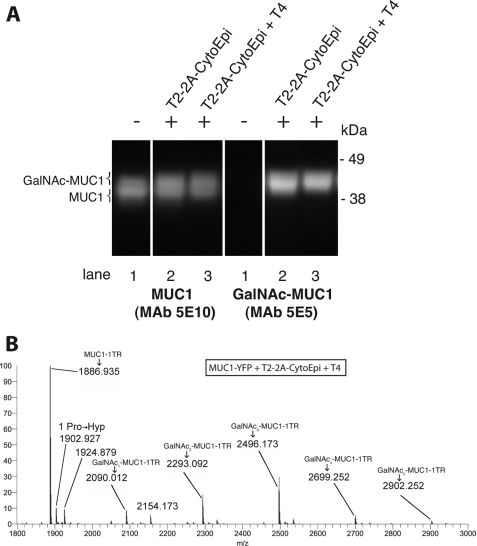
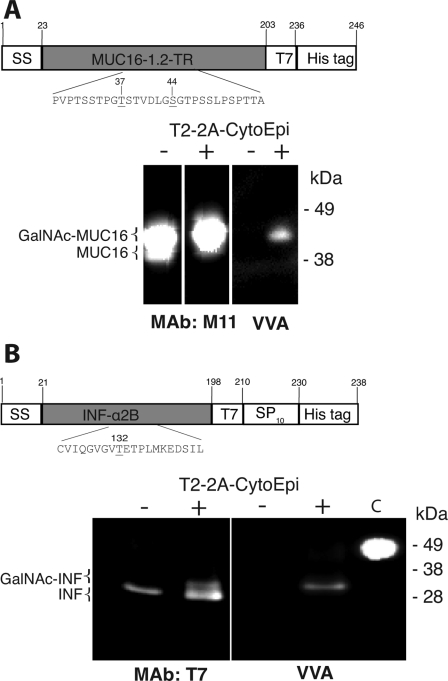
Mucin-type O-glycosylation is an important post-translational modification that confers a variety of biological properties and functions to proteins. This post-translational modification has a particularly complex and differentially regulated biosynthesis rendering prediction and control of where O-glycans are attached to proteins, and which structures are formed, difficult. Because plants are devoid of GalNAc-type O-glycosylation, we have assessed requirements for establishing human GalNAc O-glycosylation de novo in plants with the aim of developing cell systems with custom-designed O-glycosylation capacity. Transient expression of a Pseudomonas aeruginosa Glc(NAc) C4-epimerase and a human polypeptide GalNAc-transferase in leaves of Nicotiana benthamiana resulted in GalNAc O-glycosylation of co-expressed human O-glycoprotein substrates. A chimeric YFP construct containing a 3.5 tandem repeat sequence of MUC1 was glycosylated with up to three and five GalNAc residues when co-expressed with GalNAc-T2 and a combination of GalNAc-T2 and GalNAc-T4, respectively, as determined by mass spectrometry. O-Glycosylation was furthermore demonstrated on a tandem repeat of MUC16 and interferon α2b. In plants, prolines in certain classes of proteins are hydroxylated and further substituted with plant-specific O-glycosylation; unsubstituted hydroxyprolines were identified in our MUC1 construct. In summary, this study demonstrates that mammalian type O-glycosylation can be established in plants and that plants may serve as a host cell for production of recombinant O-glycoproteins with custom-designed O-glycosylation. The observed hydroxyproline modifications, however, call for additional future engineering efforts.
Figures
Similar articles
-
Toward stable genetic engineering of human O-glycosylation in plants.Plant Physiol. 2012 Sep;160(1):450-63. doi: 10.1104/pp.112.198200. Epub 2012 Jul 12. Plant Physiol. 2012. PMID: 22791304 Free PMC article.
-
Engineering of N. benthamiana L. plants for production of N-acetylgalactosamine-glycosylated proteins--towards development of a plant-based platform for production of protein therapeutics with mucin type O-glycosylation.BMC Biotechnol. 2010 Aug 24;10:62. doi: 10.1186/1472-6750-10-62. BMC Biotechnol. 2010. PMID: 20735851 Free PMC article.
-
Site-specific O-glycosylation of N-terminal serine residues by polypeptide GalNAc-transferase 2 modulates human δ-opioid receptor turnover at the plasma membrane.Cell Signal. 2018 Jan;42:184-193. doi: 10.1016/j.cellsig.2017.10.016. Epub 2017 Oct 31. Cell Signal. 2018. PMID: 29097258
-
Diverse glycosylation of MUC1 and MUC2: potential significance in tumor immunity.J Biochem. 1999 Dec;126(6):975-85. doi: 10.1093/oxfordjournals.jbchem.a022565. J Biochem. 1999. PMID: 10578046 Review.
-
Site-specific protein O-glycosylation modulates proprotein processing - deciphering specific functions of the large polypeptide GalNAc-transferase gene family.Biochim Biophys Acta. 2012 Dec;1820(12):2079-94. doi: 10.1016/j.bbagen.2012.09.014. Epub 2012 Sep 26. Biochim Biophys Acta. 2012. PMID: 23022508 Review.
Cited by
-
Toward stable genetic engineering of human O-glycosylation in plants.Plant Physiol. 2012 Sep;160(1):450-63. doi: 10.1104/pp.112.198200. Epub 2012 Jul 12. Plant Physiol. 2012. PMID: 22791304 Free PMC article.
-
A gene responsible for prolyl-hydroxylation of moss-produced recombinant human erythropoietin.Sci Rep. 2013 Oct 22;3:3019. doi: 10.1038/srep03019. Sci Rep. 2013. PMID: 24145658 Free PMC article.
-
Advanced Plant-Based Glycan Engineering.Front Bioeng Biotechnol. 2018 Jun 14;6:81. doi: 10.3389/fbioe.2018.00081. eCollection 2018. Front Bioeng Biotechnol. 2018. PMID: 29963553 Free PMC article. Review.
-
Glycosylation of Plant-Produced Immunoglobulins.Exp Suppl. 2021;112:519-543. doi: 10.1007/978-3-030-76912-3_16. Exp Suppl. 2021. PMID: 34687021
-
Two key polymorphisms in a newly discovered allele of the Vitis vinifera TPS24 gene are responsible for the production of the rotundone precursor α-guaiene.J Exp Bot. 2016 Feb;67(3):799-808. doi: 10.1093/jxb/erv491. Epub 2015 Nov 17. J Exp Bot. 2016. PMID: 26590310 Free PMC article.
References
-
- Elliott S., Lorenzini T., Asher S., Aoki K., Brankow D., Buck L., Busse L., Chang D., Fuller J., Grant J., Hernday N., Hokum M., Hu S., Knudten A., Levin N., Komorowski R., Martin F., Navarro R., Osslund T., Rogers G., Rogers N., Trail G., Egrie J. (2003) Enhancement of therapeutic protein in vivo activities through glycoengineering. Nat. Biotechnol. 21, 414–421 - PubMed
-
- Tarp M. A., Clausen H. (2008) Mucin-type O-glycosylation and its potential use in drug and vaccine development. Biochim. Biophys. Acta 1780, 546–563 - PubMed
-
- Wandall H. H., Blixt O., Tarp M. A., Pedersen J. W., Bennett E. P., Mandel U., Ragupathi G., Livingston P. O., Hollingsworth M. A., Taylor-Papadimitriou J., Burchell J., Clausen H. (2010) Cancer biomarkers defined by autoantibody signatures to aberrant O-glycopeptide epitopes. Cancer Res. 70, 1306–1313 - PMC - PubMed
-
- Xu X., Nagarajan H., Lewis N. E., Pan S., Cai Z., Liu X., Chen W., Xie M., Wang W., Hammond S., Andersen M. R., Neff N., Passarelli B., Koh W., Fan H. C., Wang J., Gui Y., Lee K. H., Betenbaugh M. J., Quake S. R., Famili I., Palsson B. O., Wang J. (2011) The genomic sequence of the Chinese hamster ovary (CHO)-K1 cell line. Nat. Biotechnol. 29, 735–741 - PMC - PubMed
-
- Gerlach J. Q., Kilcoyne M., Eaton S., Bhavanandan V., Joshi L. (2011) Noncarbohydrate-mediated interaction of lectins with plant proteins. Adv. Exp. Med. Biol. 705, 257–269 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
