

Molecular structure and function of the novel BrnT/BrnA toxin-antitoxin system of Brucella abortus
- PMID: 22334680
- PMCID: PMC3320955
- DOI: 10.1074/jbc.M111.332163
Molecular structure and function of the novel BrnT/BrnA toxin-antitoxin system of Brucella abortus
Abstract
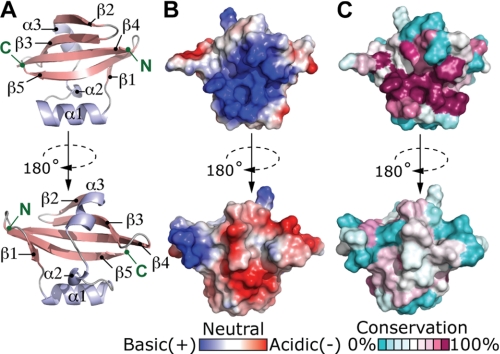
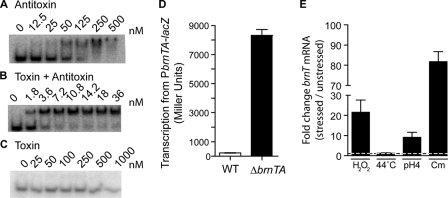
Type II toxin-antitoxin (TA) systems are expressed from two-gene operons that encode a cytoplasmic protein toxin and its cognate protein antitoxin. These gene cassettes are often present in multiple copies on bacterial chromosomes, where they have been reported to regulate stress adaptation and persistence during antimicrobial treatment. We have identified a novel type II TA cassette in the intracellular pathogen Brucella abortus that consists of the toxin gene, brnT, and its antitoxin, brnA. BrnT is coexpressed and forms a 2:2 tetrameric complex with BrnA, which neutralizes BrnT toxicity. The BrnT(2)-BrnA(2) tetramer binds its own promoter via BrnA, and autorepresses its expression; its transcription is strongly induced in B. abortus by various stressors encountered by the bacterial cell during infection of a mammalian host. Although highly divergent at the primary sequence level, an atomic resolution (1.1 Å) crystal structure of BrnT reveals a secondary topology related to the RelE family of type II ribonuclease toxins. However, overall tertiary structural homology to other RelE family toxins is low. A functional characterization of BrnT by site-directed mutagenesis demonstrates a correspondence between its in vitro activity as a ribonuclease and control of bacteriostasis in vivo. We further present an analysis of the conserved and variable features of structure required for RNA scission in BrnT and the RelE toxin family. This structural investigation informs a model of the RelE-fold as an evolutionarily flexible scaffold that has been selected to bind structurally disparate antitoxins, and exhibit distinct toxin activities including RNA scission and DNA gyrase inhibition.
Figures






Similar articles
-
Crystal Structure of VapBC-1 from Nontypeable Haemophilus influenzae and the Effect of PIN Domain Mutations on Survival during Infection.J Bacteriol. 2019 May 22;201(12):e00026-19. doi: 10.1128/JB.00026-19. Print 2019 Jun 15. J Bacteriol. 2019. PMID: 30936373 Free PMC article.
-
VapC from the leptospiral VapBC toxin-antitoxin module displays ribonuclease activity on the initiator tRNA.PLoS One. 2014 Jul 21;9(7):e101678. doi: 10.1371/journal.pone.0101678. eCollection 2014. PLoS One. 2014. PMID: 25047537 Free PMC article.
-
Functional details of the Mycobacterium tuberculosis VapBC26 toxin-antitoxin system based on a structural study: insights into unique binding and antibiotic peptides.Nucleic Acids Res. 2017 Aug 21;45(14):8564-8580. doi: 10.1093/nar/gkx489. Nucleic Acids Res. 2017. PMID: 28575388 Free PMC article.
-
Structure and function of bacterial kid-kis and related toxin-antitoxin systems.Protein Pept Lett. 2007;14(2):113-24. doi: 10.2174/092986607779816096. Protein Pept Lett. 2007. PMID: 17305597 Review.
-
Toxins, Targets, and Triggers: An Overview of Toxin-Antitoxin Biology.Mol Cell. 2018 Jun 7;70(5):768-784. doi: 10.1016/j.molcel.2018.01.003. Epub 2018 Feb 3. Mol Cell. 2018. PMID: 29398446 Review.
Cited by
-
Trimethoprim resistance in surface and wastewater is mediated by contrasting variants of the dfrB gene.ISME J. 2023 Sep;17(9):1455-1466. doi: 10.1038/s41396-023-01460-7. Epub 2023 Jun 27. ISME J. 2023. PMID: 37369703 Free PMC article.
-
Enhanced transformation efficiency in Treponema denticola enabled by SyngenicDNA-based plasmids lacking restriction-modification target motifs.Mol Oral Microbiol. 2023 Dec;38(6):455-470. doi: 10.1111/omi.12441. Epub 2023 Oct 25. Mol Oral Microbiol. 2023. PMID: 37880921 Free PMC article.
-
The endogenous Coxiella burnetii plasmid encodes a functional toxin-antitoxin system.Mol Microbiol. 2022 Dec;118(6):744-764. doi: 10.1111/mmi.15001. Epub 2022 Nov 28. Mol Microbiol. 2022. PMID: 36385554 Free PMC article.
-
Toxin-antitoxin systems as multilevel interaction systems.Toxins (Basel). 2014 Jan 10;6(1):304-24. doi: 10.3390/toxins6010304. Toxins (Basel). 2014. PMID: 24434905 Free PMC article. Review.
-
CoCoNuTs are a diverse subclass of Type IV restriction systems predicted to target RNA.Elife. 2024 May 13;13:RP94800. doi: 10.7554/eLife.94800. Elife. 2024. PMID: 38739430 Free PMC article.
References
-
- Anderson T. D., Meador V. P., Cheville N. F. (1986) Pathogenesis of placentitis in the goat inoculated with Brucella abortus. I. Gross and histologic lesions. Vet. Pathol. 23, 219–226 - PubMed
-
- Moreno E., Moriyon I. (2006) in The Prokaryotes: A Handbook on the Biology of Bacteria (Dworkin M., Falkow S., Rosenberg E., Schleifer K. H., Stackebrandt E., eds) 3rd Ed., pp. 315–456, Springer, New York
-
- Jiang X., Leonard B., Benson R., Baldwin C. L. (1993) Macrophage control of Brucella abortus. Role of reactive oxygen intermediates and nitric oxide. Cell. Immunol. 151, 309–319 - PubMed
-
- Köhler S., Michaux-Charachon S., Porte F., Ramuz M., Liautard J. P. (2003) What is the nature of the replicative niche of a stealthy bug named Brucella? Trends Microbiol. 11, 215–219 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
