

Nuclear-localized and deregulated calcium- and calmodulin-dependent protein kinase activates rhizobial and mycorrhizal responses in Lotus japonicus
- PMID: 22337918
- PMCID: PMC3315248
- DOI: 10.1105/tpc.111.091827
Nuclear-localized and deregulated calcium- and calmodulin-dependent protein kinase activates rhizobial and mycorrhizal responses in Lotus japonicus
Abstract
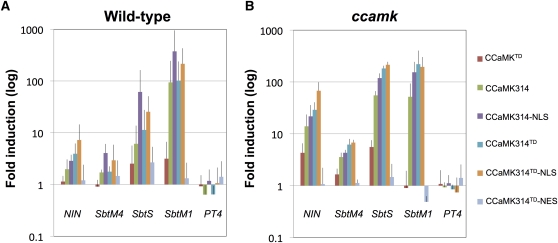
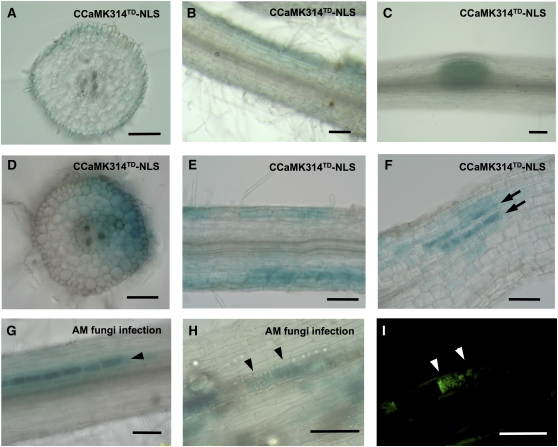
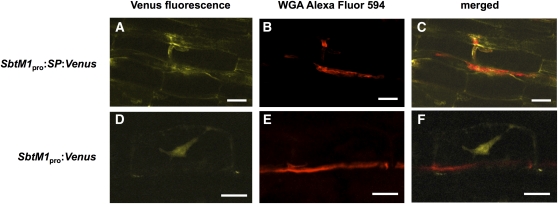
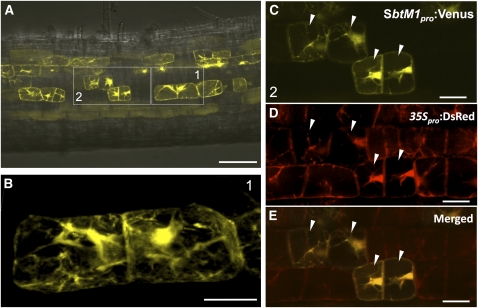
The common symbiosis pathway is at the core of symbiosis signaling between plants and soil microbes. In this pathway, calcium- and calmodulin-dependent protein kinase (CCaMK) plays a crucial role in integrating the signals both in arbuscular mycorrhizal symbiosis (AMS) and in root nodule symbiosis (RNS). However, the molecular mechanism by which CCaMK coordinates AMS and RNS is largely unknown. Here, we report that the gain-of-function (GOF) variants of CCaMK without the regulatory domains activate both AMS and RNS signaling pathways in the absence of symbiotic partners. This activation requires nuclear localization of CCaMK. Enforced nuclear localization of the GOF-CCaMK variants by fusion with a canonical nuclear localization signal enhances signaling activity of AMS and RNS. The GOF-CCaMK variant triggers formation of a structure similar to the prepenetration apparatus, which guides infection of arbuscular mycorrhizal fungi to host root cells. In addition, the GOF-CCaMK variants without the regulatory domains partly restore AMS but fail to support rhizobial infection in ccamk mutants. These data indicate that AMS, the more ancient type of symbiosis, can be mainly regulated by the kinase activity of CCaMK, whereas RNS, which evolved more recently, requires complex regulation performed by the regulatory domains of CCaMK.
Figures
References
-
- Ané J.M., et al. (2004). Medicago truncatula DMI1 required for bacterial and fungal symbioses in legumes. Science 303: 1364–1367 - PubMed
-
- Ardourel M., Demont N., Debellé F., Maillet F., de Billy F., Promé J.C., Dénarié J., Truchet G. (1994). Rhizobium meliloti lipooligosaccharide nodulation factors: Different structural requirements for bacterial entry into target root hair cells and induction of plant symbiotic developmental responses. Plant Cell 6: 1357–1374 - PMC - PubMed
-
- Banba M., Gutjahr C., Miyao A., Hirochika H., Paszkowski U., Kouchi H., Imaizumi-Anraku H. (2008). Divergence of evolutionary ways among common sym genes: CASTOR and CCaMK show functional conservation between two symbiosis systems and constitute the root of a common signaling pathway. Plant Cell Physiol. 49: 1659–1671 - PubMed
-
- Bonfante P., Perotto S. (1995). Strategies of arbuscular mycorrhizal fungi when infecting host plants. New Phytol. 130: 3–21
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
