

Neurological diseases as primary gliopathies: a reassessment of neurocentrism
- PMID: 22339481
- PMCID: PMC3320215
- DOI: 10.1042/AN20120010
Neurological diseases as primary gliopathies: a reassessment of neurocentrism
Abstract
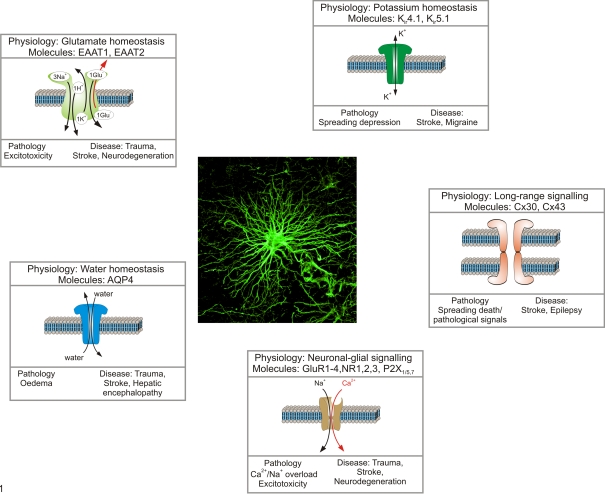
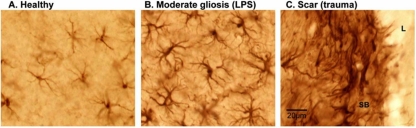
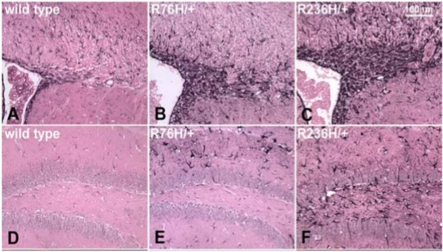
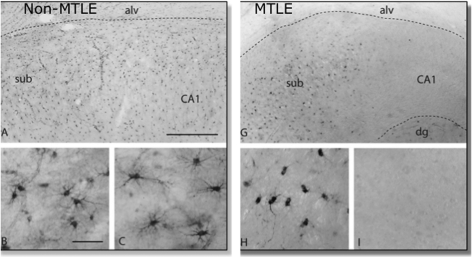
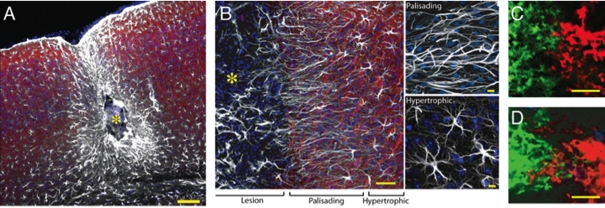
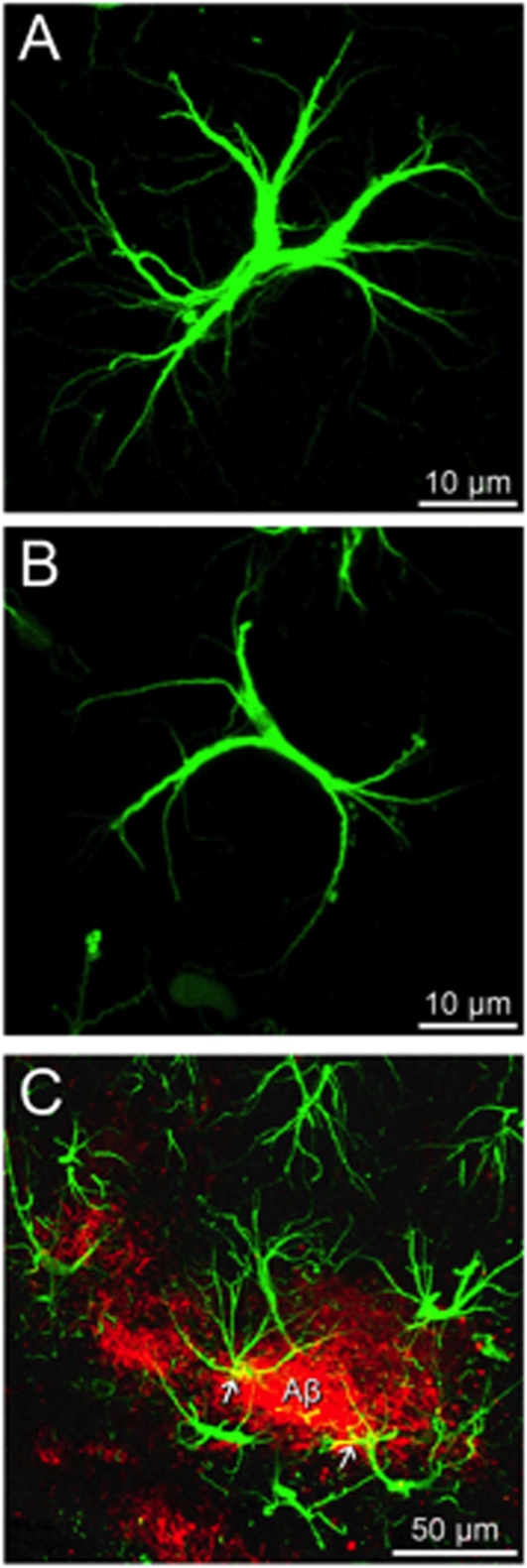
Diseases of the human brain are almost universally attributed to malfunction or loss of nerve cells. However, a considerable amount of work has, during the last decade, expanded our view on the role of astrocytes in CNS (central nervous system), and this analysis suggests that astrocytes contribute to both initiation and propagation of many (if not all) neurological diseases. Astrocytes provide metabolic and trophic support to neurons and oligodendrocytes. Here, we shall endeavour a broad overviewing of the progress in the field and forward the idea that loss of homoeostatic astroglial function leads to an acute loss of neurons in the setting of acute insults such as ischaemia, whereas more subtle dysfunction of astrocytes over periods of months to years contributes to epilepsy and to progressive loss of neurons in neurodegenerative diseases. The majority of therapeutic drugs currently in clinical use target neuronal receptors, channels or transporters. Future therapeutic efforts may benefit by a stronger focus on the supportive homoeostatic functions of astrocytes.
Figures
References
-
- Alexander WS. Progressive fibrinoid degeneration of fibrillary astrocytes associated with mental retardation in a hydrocephalic infant. Brain. 1949;72:373–381. - PubMed
-
- Alvarez-Buylla A, Garcia-Verdugo JM, Tramontin AD. A unified hypothesis on the lineage of neural stem cells. Nat Rev Neurosci. 2001;2:287–293. - PubMed
-
- Alzheimer A, Nissl F, Alzheimer A. Beiträge zur Kenntnis der pathologischen Neuroglia und ihrer Beziehungen zu den Abbauvorgängen im Nervengewebe. Histologische und Histopathologische Arbeiten über die Grosshirnrinde mit besonderer Berücksichtigung der pathologischen Anatomie der Geisteskrankheiten. Verlag von Gustav Fischer Jena. 1910:401–562.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical