

Biologic scaffolds composed of central nervous system extracellular matrix
- PMID: 22341938
- PMCID: PMC3516286
- DOI: 10.1016/j.biomaterials.2012.01.044
Biologic scaffolds composed of central nervous system extracellular matrix
Abstract
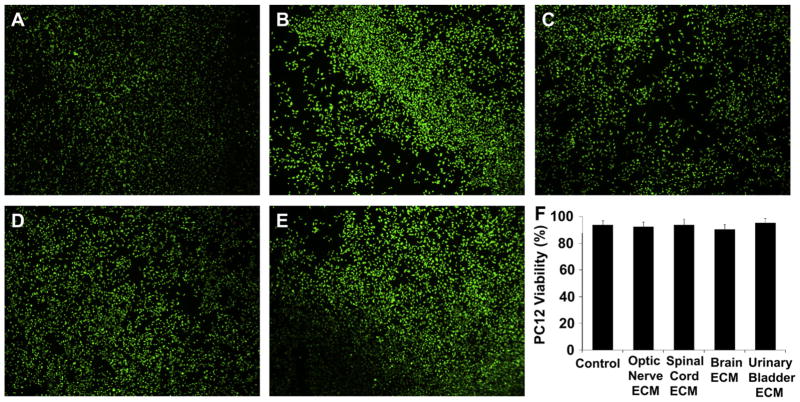
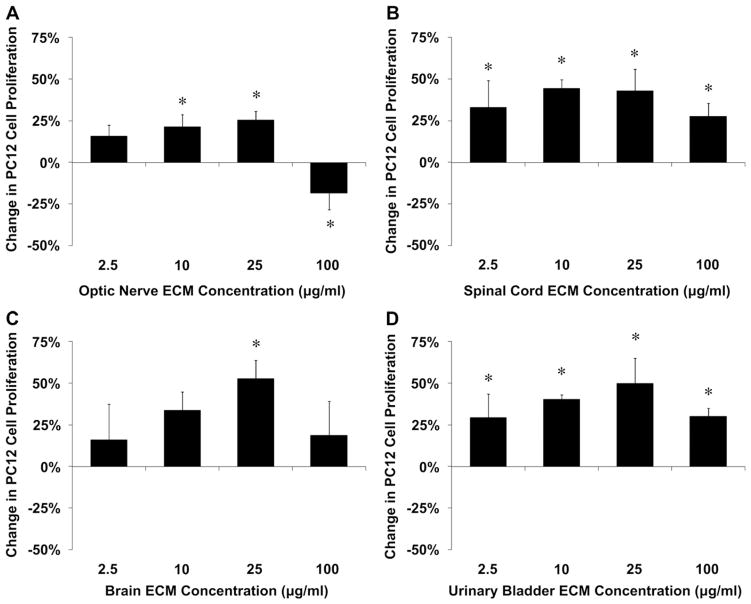
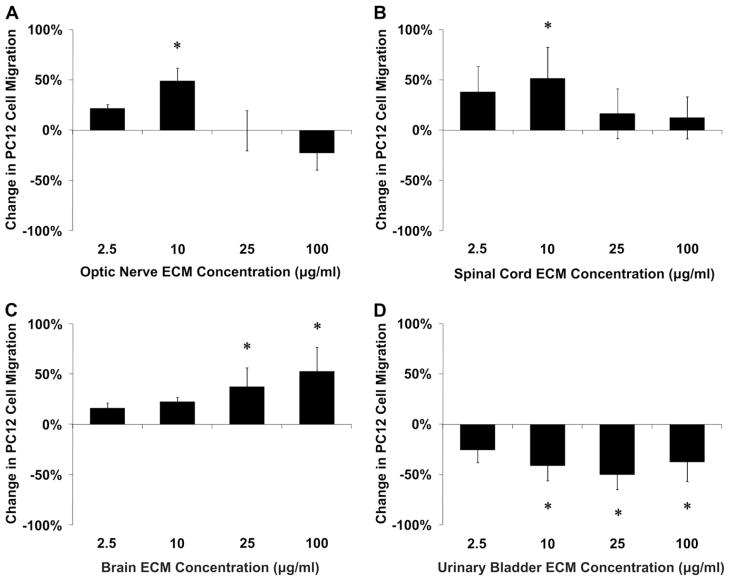
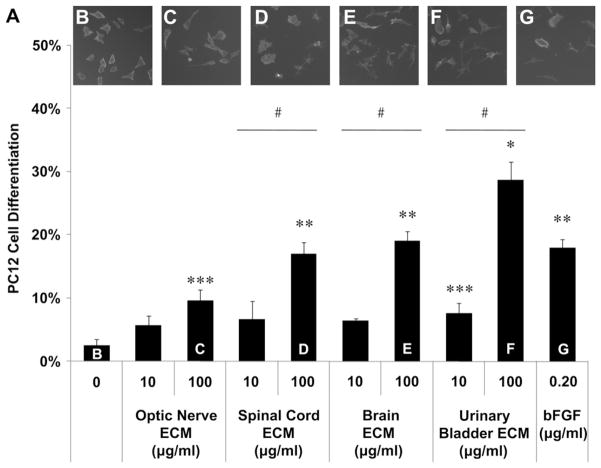
Acellular biologic scaffolds are commonly used to facilitate the constructive remodeling of three of the four traditional tissue types: connective, epithelial, and muscle tissues. However, the application of extracellular matrix (ECM) scaffolds to neural tissue has been limited, particularly in the central nervous system (CNS) where intrinsic regenerative potential is low. The ability of decellularized liver, lung, muscle, and other tissues to support tissue-specific cell phenotype and function suggests that CNS-derived biologic scaffolds may help to overcome barriers to mammalian CNS repair. A method was developed to create CNS ECM scaffolds from porcine optic nerve, spinal cord, and brain, with decellularization verified against established criteria. CNS ECM scaffolds retained neurosupportive proteins and growth factors and, when tested with the PC12 cell line in vitro, were cytocompatible and stimulated proliferation, migration, and differentiation. Urinary bladder ECM (a non-CNS ECM scaffold) was also cytocompatible and stimulated PC12 proliferation but inhibited migration rather than acting as a chemoattractant over the same concentration range while inducing greater rates of PC12 differentiation compared to CNS ECM. These results suggest that CNS ECM may provide tissue-specific advantages in CNS regenerative medicine applications and that ECM scaffolds in general may aid functional recovery after CNS injury.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
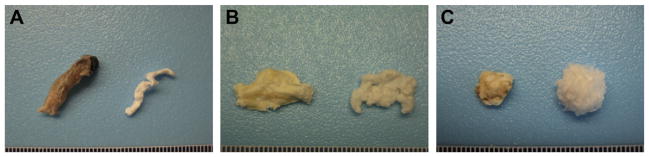
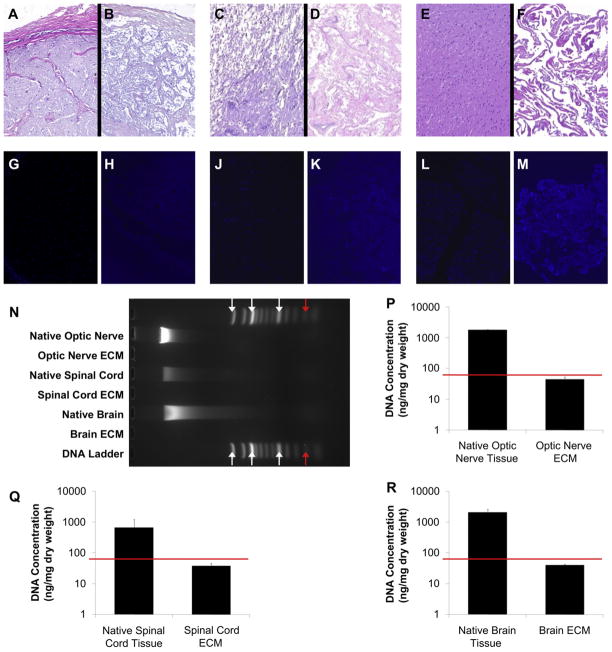
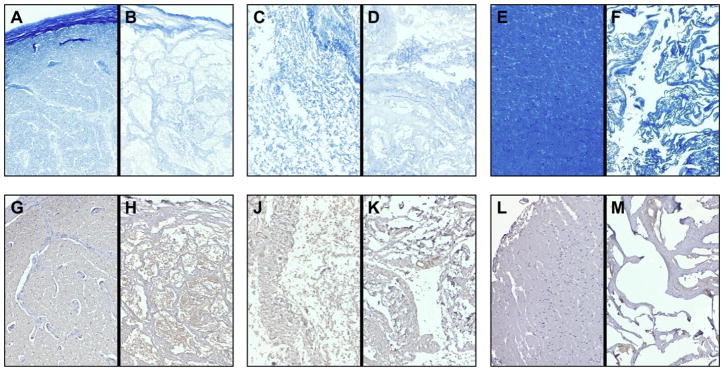
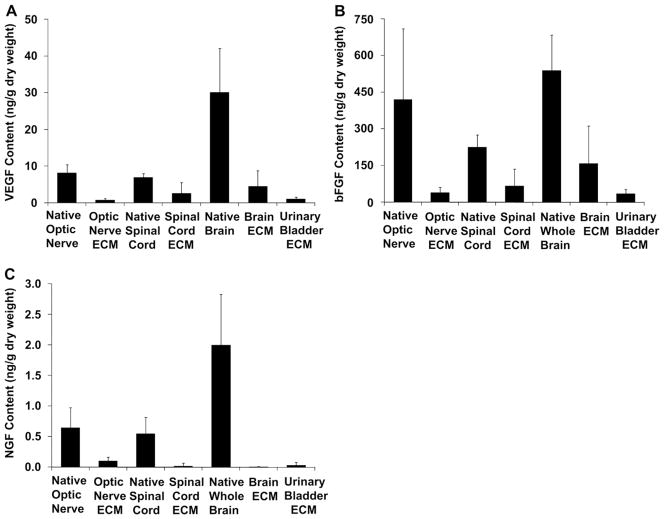
Similar articles
-
Effects of biologic scaffolds on human stem cells and implications for CNS tissue engineering.Tissue Eng Part A. 2014 Jan;20(1-2):313-23. doi: 10.1089/ten.TEA.2013.0186. Epub 2013 Oct 10. Tissue Eng Part A. 2014. PMID: 24004192 Free PMC article.
-
Biologic scaffold composed of skeletal muscle extracellular matrix.Biomaterials. 2012 Apr;33(10):2916-25. doi: 10.1016/j.biomaterials.2011.12.055. Epub 2012 Jan 20. Biomaterials. 2012. PMID: 22264525 Free PMC article.
-
Targeted proteomics effectively quantifies differences between native lung and detergent-decellularized lung extracellular matrices.Acta Biomater. 2016 Dec;46:91-100. doi: 10.1016/j.actbio.2016.09.043. Epub 2016 Sep 29. Acta Biomater. 2016. PMID: 27693690 Free PMC article.
-
Biologic scaffold for CNS repair.Regen Med. 2014 May;9(3):367-83. doi: 10.2217/rme.14.9. Regen Med. 2014. PMID: 24935046 Review.
-
Methods of tissue decellularization used for preparation of biologic scaffolds and in vivo relevance.Methods. 2015 Aug;84:25-34. doi: 10.1016/j.ymeth.2015.03.005. Epub 2015 Mar 16. Methods. 2015. PMID: 25791470 Review.
Cited by
-
Regeneration strategies after the adult mammalian central nervous system injury-biomaterials.Regen Biomater. 2016 Jun;3(2):115-22. doi: 10.1093/rb/rbw004. Epub 2016 Mar 8. Regen Biomater. 2016. PMID: 27047678 Free PMC article. Review.
-
Acquisition of Visual Perception in Blind Adults Using the BrainPort Artificial Vision Device.Am J Occup Ther. 2015 Jan-Feb;69(1):6901290010p1-8. doi: 10.5014/ajot.2015.011809. Am J Occup Ther. 2015. PMID: 25553750 Free PMC article.
-
Mechanotransduction: Exploring New Therapeutic Avenues in Central Nervous System Pathology.Front Neurosci. 2022 Apr 28;16:861613. doi: 10.3389/fnins.2022.861613. eCollection 2022. Front Neurosci. 2022. PMID: 35573316 Free PMC article. Review.
-
A roadmap for promoting endogenous in situ tissue restoration using inductive bioscaffolds after acute brain injury.Brain Res Bull. 2019 Aug;150:136-149. doi: 10.1016/j.brainresbull.2019.05.013. Epub 2019 May 22. Brain Res Bull. 2019. PMID: 31128250 Free PMC article. Review.
-
Neuroprotective Effects of Collagen-Glycosaminoglycan Matrix Implantation following Surgical Brain Injury.Mediators Inflamm. 2019 Jan 27;2019:6848943. doi: 10.1155/2019/6848943. eCollection 2019. Mediators Inflamm. 2019. PMID: 30809107 Free PMC article.
References
-
- Vorotnikova E, McIntosh D, Dewilde A, Zhang J, Reing JE, Zhang L, et al. Extracellular matrix-derived products modulate endothelial and progenitor cell migration and proliferation in vitro and stimulate regenerative healing in vivo. Matrix Biol. 2010;29:690–700. - PubMed
-
- Cortiella J, Niles J, Cantu A, Brettler A, Pham A, Vargas G, et al. Influence of acellular natural lung matrix on murine embryonic stem cell differentiation and tissue formation. Tissue Eng Part A. 2010;16:2565–80. - PubMed
-
- Ott HC, Clippinger B, Conrad C, Schuetz C, Pomerantseva I, Ikonomou L, et al. Regeneration and orthotopic transplantation of a bioartificial lung. Nat Med. 2010;16:927–33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
