

Defining "epileptogenesis" and identifying "antiepileptogenic targets" in animal models of acquired temporal lobe epilepsy is not as simple as it might seem
- PMID: 22342985
- PMCID: PMC3398197
- DOI: 10.1016/j.neuropharm.2012.01.022
Defining "epileptogenesis" and identifying "antiepileptogenic targets" in animal models of acquired temporal lobe epilepsy is not as simple as it might seem
Abstract
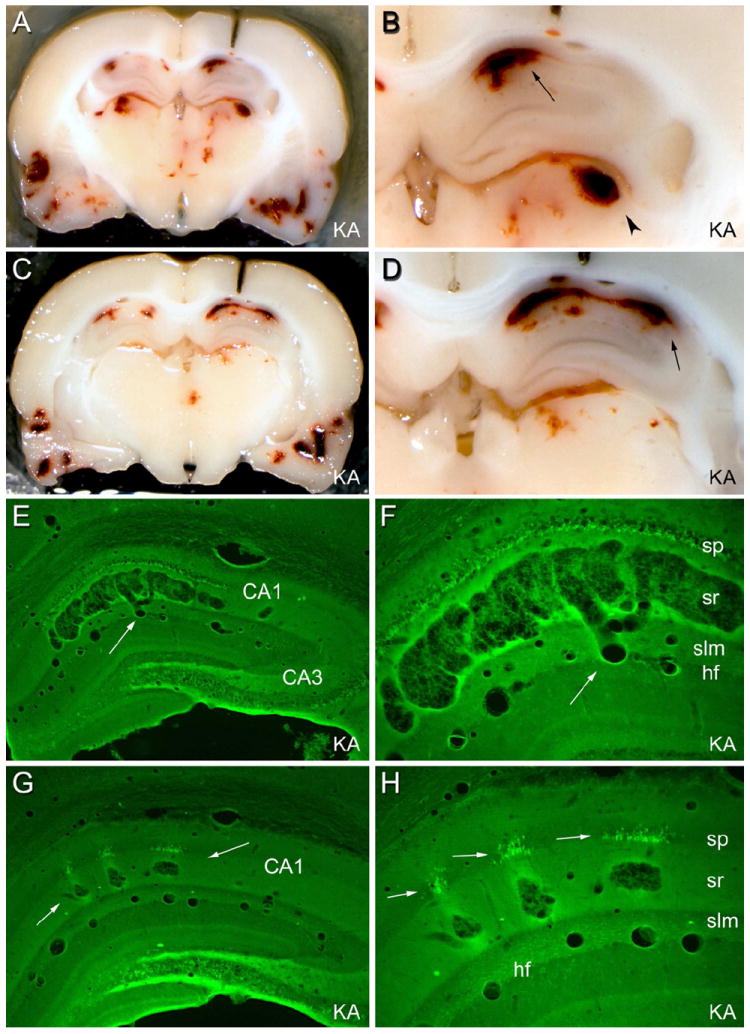
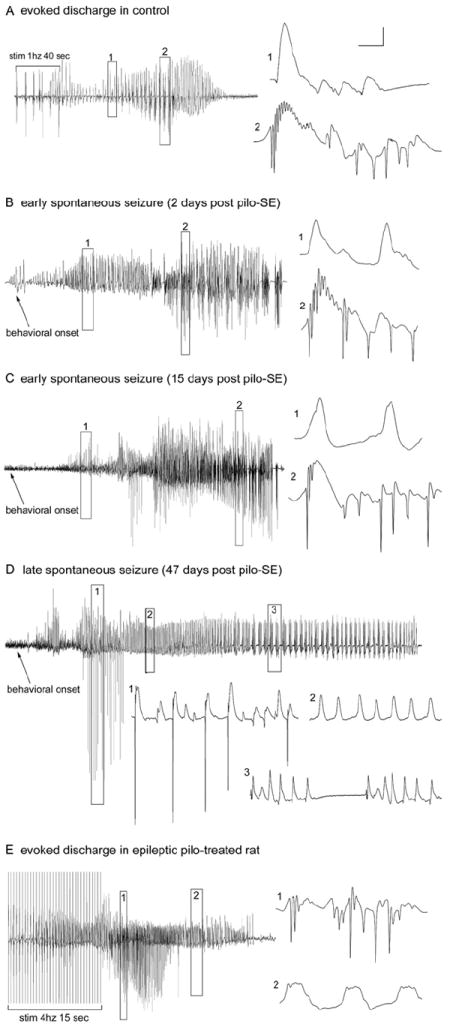
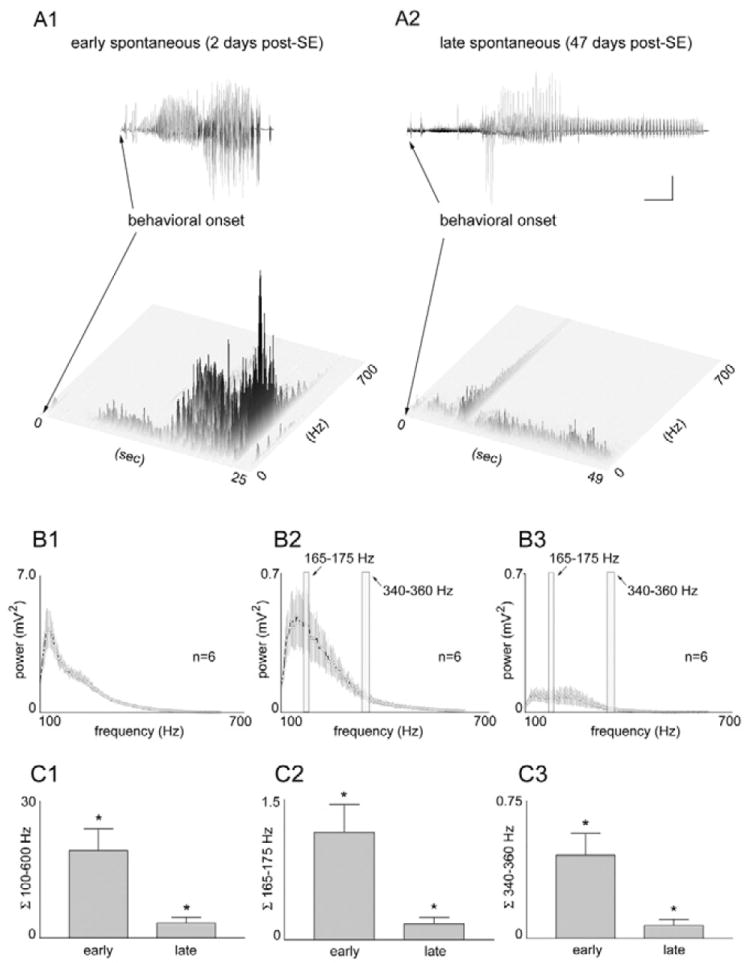
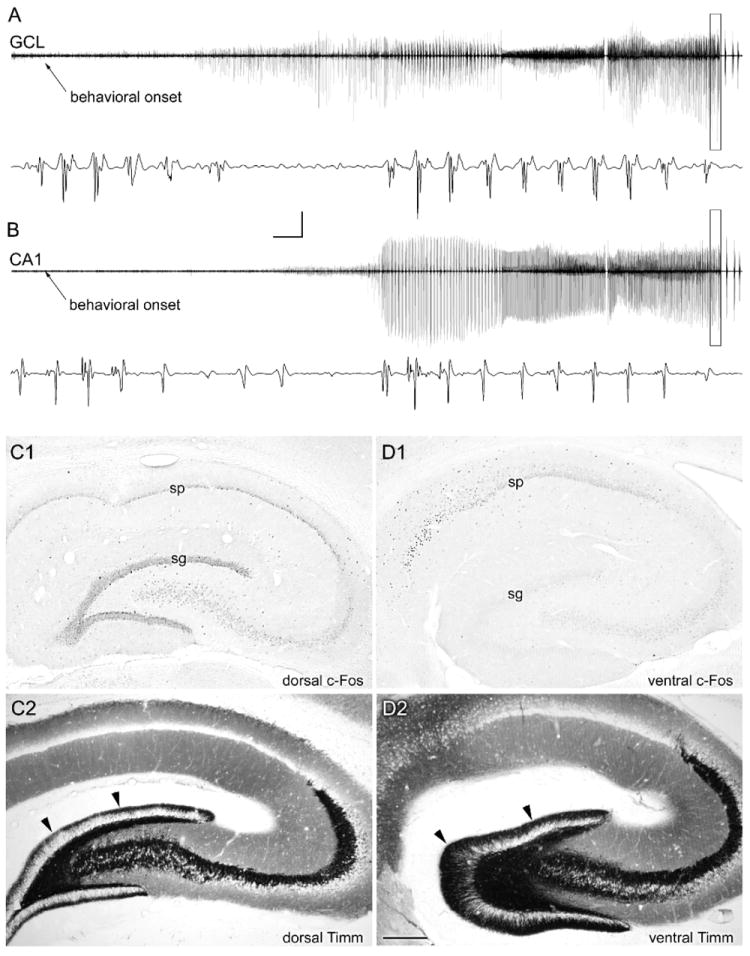
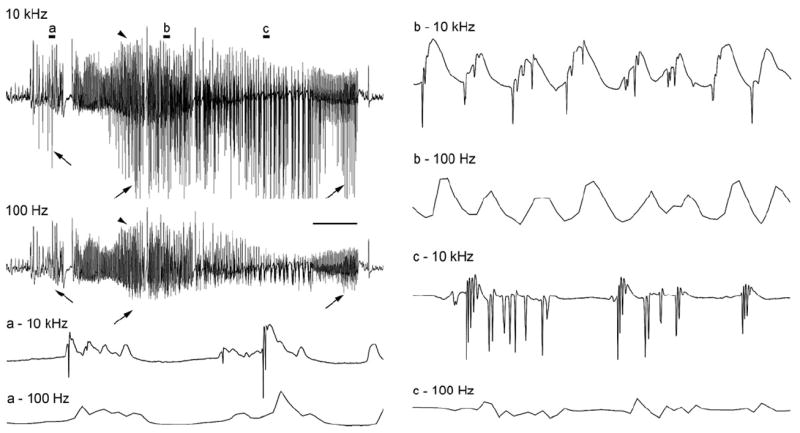
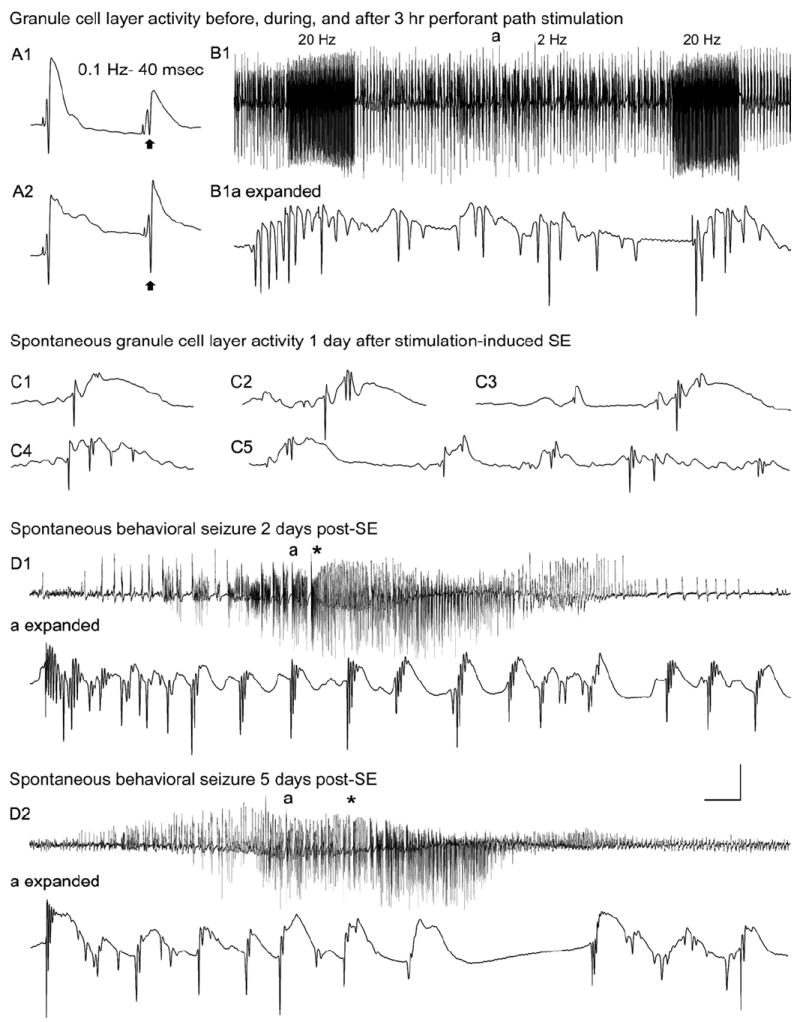
The "latent period" between brain injury and clinical epilepsy is widely regarded to be a seizure-free, pre-epileptic state during which a time-consuming cascade of molecular events and structural changes gradually mediates the process of "epileptogenesis." The concept of the "latent period" as the duration of "epileptogenesis" implies that epilepsy is not an immediate result of brain injury, and that anti-epileptogenic strategies need to target delayed secondary mechanisms that develop sometime after an initial injury. However, depth recordings made directly from the dentate granule cell layers in awake rats after convulsive status epilepticus-induced injury have now shown that whenever perforant pathway stimulation-induced status epilepticus produces extensive hilar neuron loss and entorhinal cortical injury, hyperexcitable granule cells immediately generate spontaneous epileptiform discharges and focal or generalized behavioral seizures. This indicates that hippocampal injury caused by convulsive status epilepticus is immediately epileptogenic and that hippocampal epileptogenesis requires no delayed secondary mechanism. When latent periods do exist after injury, we hypothesize that less extensive cell loss causes an extended period during which initially subclinical focal seizures gradually increase in duration to produce the first clinical seizure. Thus, the "latent period" is suggested to be a state of "epileptic maturation," rather than a prolonged period of "epileptogenesis," and therefore the antiepileptogenic therapeutic window may only remain open during the first week after injury, when some delayed cell death may still be preventable. Following the perhaps unavoidable development of the first focal seizures ("epileptogenesis"), the most fruitful therapeutic strategy may be to interrupt the process of "epileptic maturation," thereby keeping focal seizures focal. This article is part of the Special Issue entitled 'New Targets and Approaches to the Treatment of Epilepsy'.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
References
-
- Andersen P, Holmqvist B, Voorhoeve PE. Entorhinal activation of dentate granule cells. Acta Physiol Scand. 1966;66:448–460. - PubMed
-
- Ben-Ari Y. Limbic seizure and brain damage produced by kainic acid: mechanisms and relevance to human temporal lobe epilepsy. Neuroscience. 1985;14:375–403. - PubMed
-
- Blümcke I, Pauli E, Clusmann H, Schramm J, Becker A, Elger C, Merschhemke M, Meencke HJ, Lehmann T, von Deimling A, Scheiwe C, Zentner J, Volk B, Romstöck J, Stefan H, Hildebrandt M. A new clinico-pathological classification system for mesial temporal sclerosis. Acta Neuropathol. 2007;113:235–244. - PMC - PubMed
-
- Bower MR, Buckmaster PS. Changes in granule cell firing rates precede locally recorded spontaneous seizures by minutes in an animal model of temporal lobe epilepsy. J Neurophysiol. 2008;99:2431–2442. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
