

Two cation transporters Ena1 and Nha1 cooperatively modulate ion homeostasis, antifungal drug resistance, and virulence of Cryptococcus neoformans via the HOG pathway
- PMID: 22343280
- PMCID: PMC3319253
- DOI: 10.1016/j.fgb.2012.02.001
Two cation transporters Ena1 and Nha1 cooperatively modulate ion homeostasis, antifungal drug resistance, and virulence of Cryptococcus neoformans via the HOG pathway
Abstract
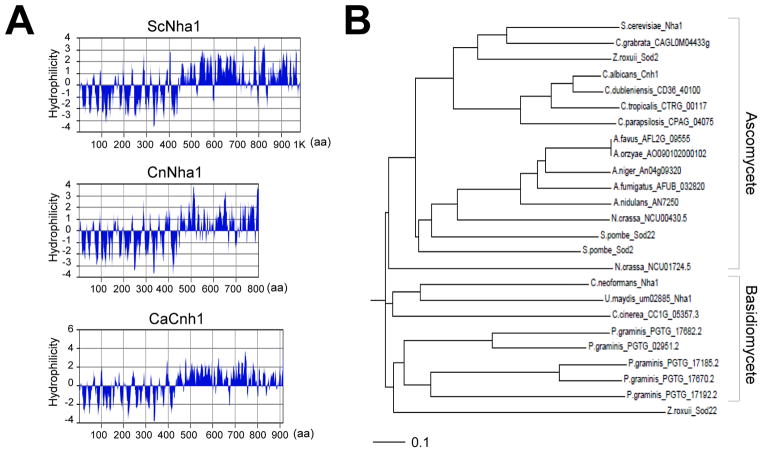
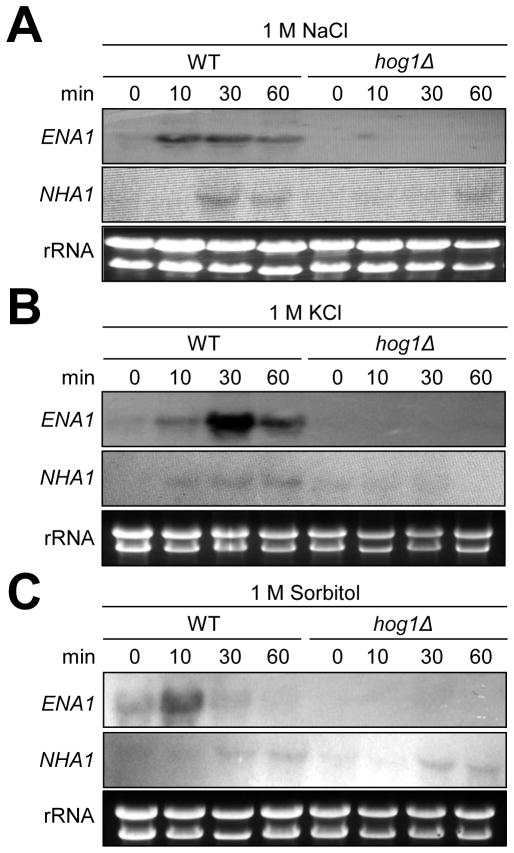
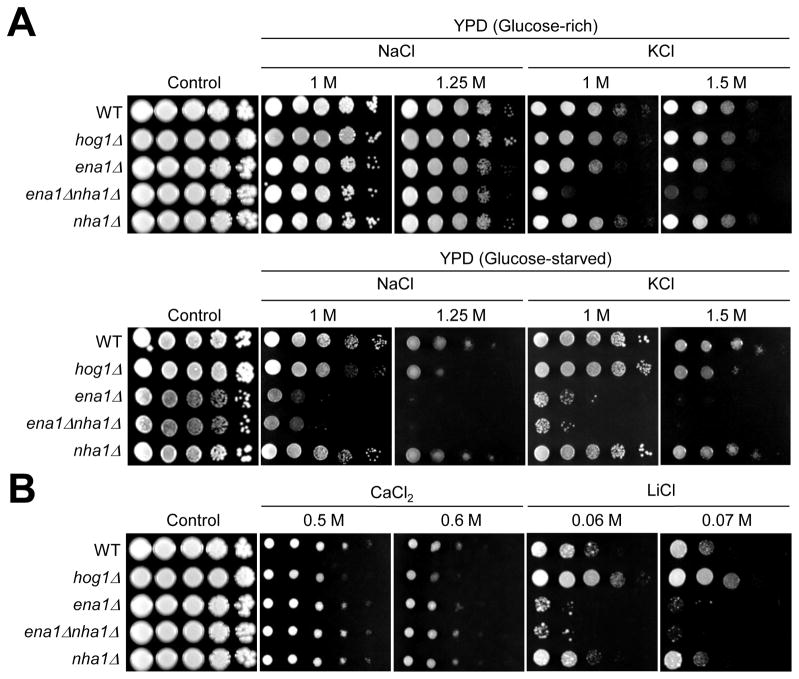
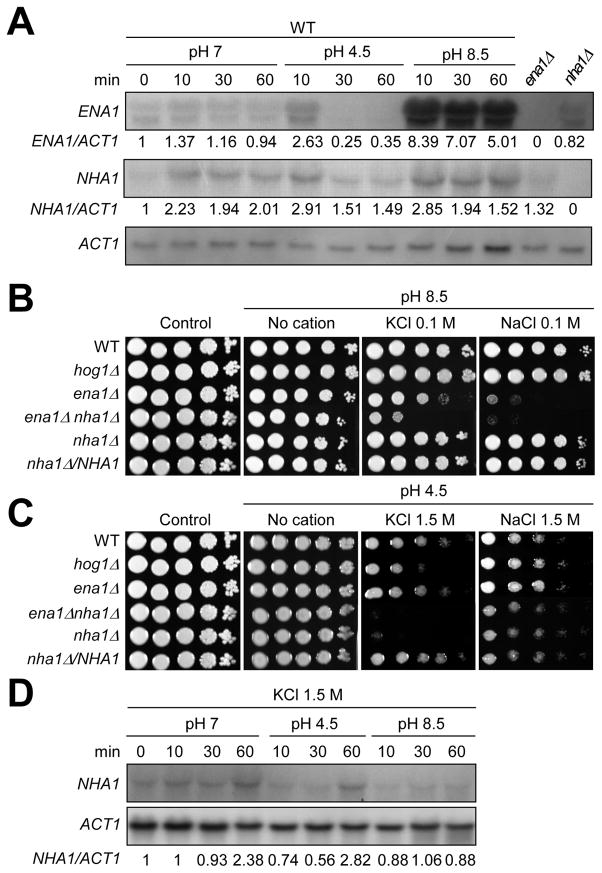
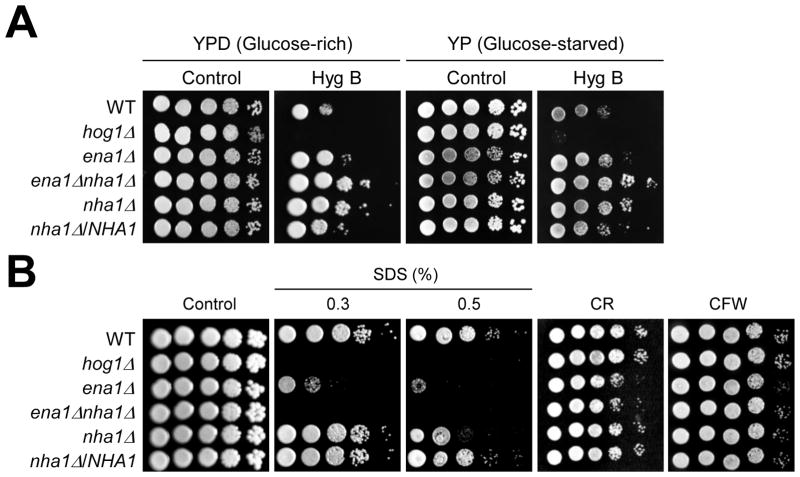
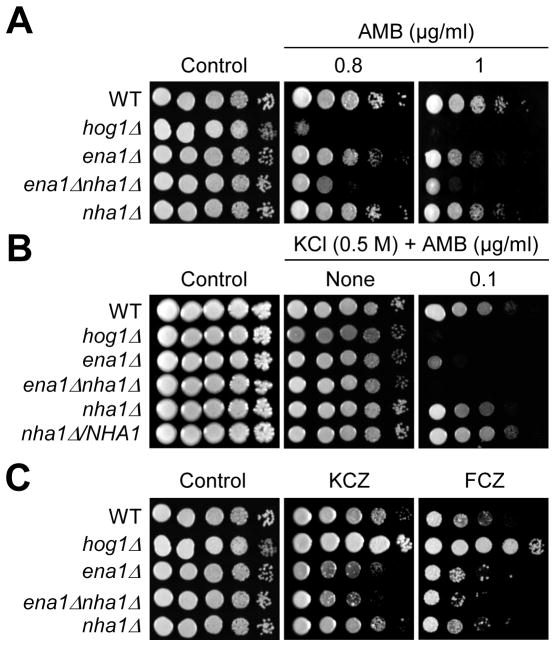
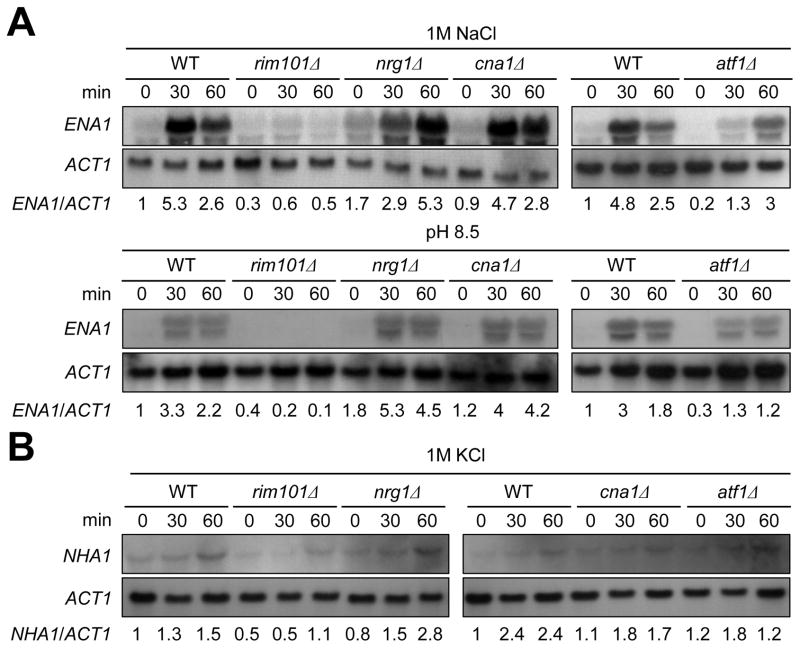
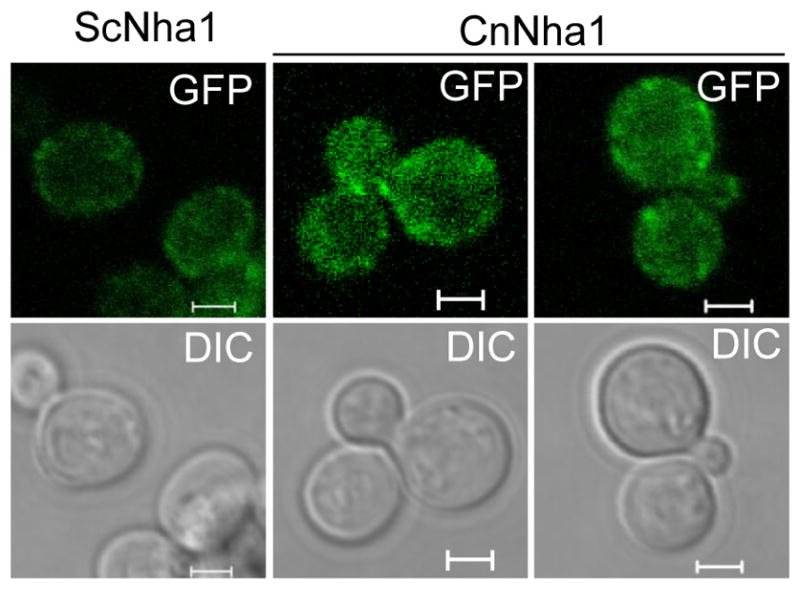
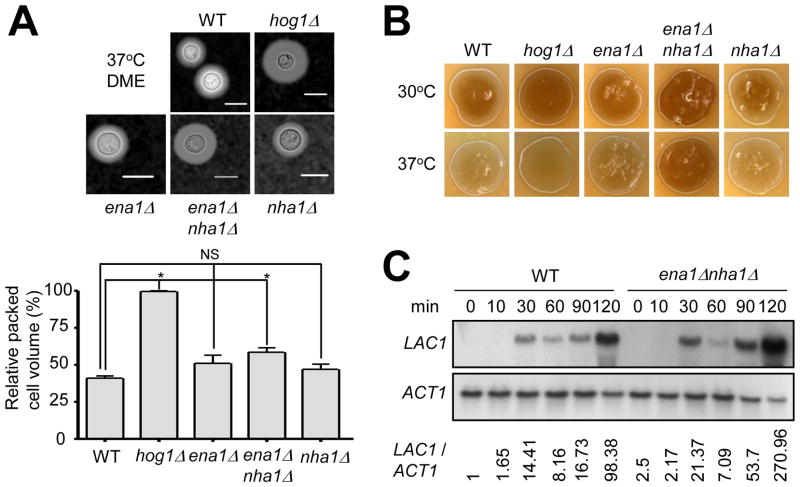
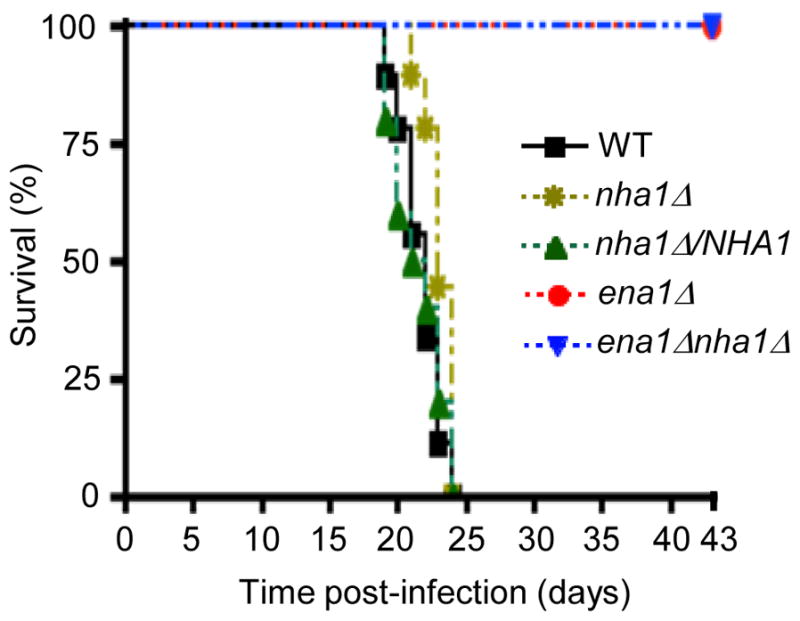
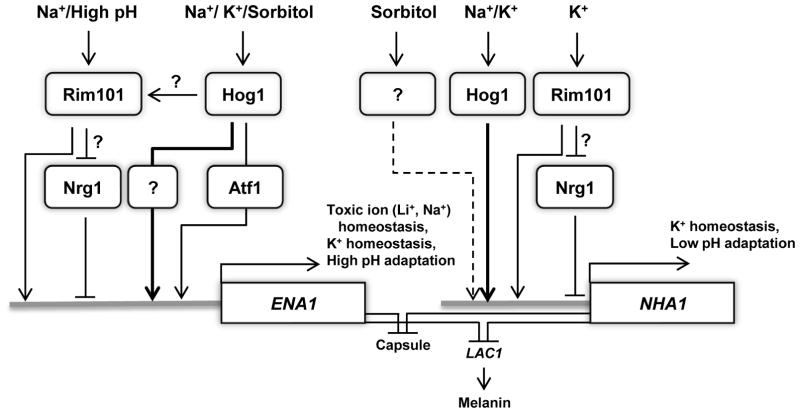
Maintenance of cation homeostasis is essential for survival of all living organisms in their biological niches. It is also important for the survival of human pathogenic fungi in the host, where cation concentrations and pH will vary depending on different anatomical sites. However, the exact role of diverse cation transporters and ion channels in virulence of fungal pathogens remains elusive. In this study we functionally characterized ENA1 and NHA1, encoding a putative Na(+)/ATPase and Na(+)/H(+) antiporter, respectively, in Cryptococcus neoformans, a basidiomycete fungal pathogen which causes fatal meningoencephalitis. Expression of NHA1 and ENA1 is induced in response to salt and osmotic shock mainly in a Hog1-dependent manner. Phenotypic analysis of the ena1Δ, nha1Δ, and ena1Δnha1Δ mutants revealed that Ena1 controls cellular levels of toxic cations, such as Na(+) and Li(+) whereas both Ena1 and Nha1 are important for controlling less toxic K(+) ions. Under alkaline conditions, Ena1 was highly induced and required for growth in the presence of low levels of Na(+) or K(+) salt and Nha1 played a role in survival under K(+) stress. In contrast, Nha1, but not Ena1, was essential for survival at acidic conditions (pH 4.5) under high K(+) stress. In addition, Ena1 and Nha1 were required for maintenance of plasma membrane potential and stability, which appeared to modulate antifungal drug susceptibility. Perturbation of ENA1 and NHA1 enhanced capsule production and melanin synthesis. However, Nha1 was dispensable for virulence of C. neoformans although Ena1 was essential. In conclusion, Ena1 and Nha1 play redundant and discrete roles in cation homeostasis, pH regulation, membrane potential, and virulence in C. neoformans, suggesting that these transporters could be novel antifungal drug targets for treatment of cryptococcosis.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
A flucytosine-responsive Mbp1/Swi4-like protein, Mbs1, plays pleiotropic roles in antifungal drug resistance, stress response, and virulence of Cryptococcus neoformans.Eukaryot Cell. 2012 Jan;11(1):53-67. doi: 10.1128/EC.05236-11. Epub 2011 Nov 11. Eukaryot Cell. 2012. PMID: 22080454 Free PMC article.
-
Identification of ENA1 as a virulence gene of the human pathogenic fungus Cryptococcus neoformans through signature-tagged insertional mutagenesis.Eukaryot Cell. 2009 Mar;8(3):315-26. doi: 10.1128/EC.00375-08. Epub 2009 Jan 16. Eukaryot Cell. 2009. PMID: 19151325 Free PMC article.
-
Lipid Flippase Subunit Cdc50 Mediates Drug Resistance and Virulence in Cryptococcus neoformans.mBio. 2016 May 10;7(3):e00478-16. doi: 10.1128/mBio.00478-16. mBio. 2016. PMID: 27165800 Free PMC article.
-
Monovalent cation transporters at the plasma membrane in yeasts.Yeast. 2019 Apr;36(4):177-193. doi: 10.1002/yea.3355. Epub 2018 Oct 3. Yeast. 2019. PMID: 30193006 Review.
-
Induction of signal transduction pathways related to the pathogenicity of Cryptococcus neoformans in the host environment.Drug Discov Ther. 2019;13(4):177-182. doi: 10.5582/ddt.2019.01047. Drug Discov Ther. 2019. PMID: 31534068 Review.
Cited by
-
Unique roles of the unfolded protein response pathway in fungal development and differentiation.Sci Rep. 2016 Sep 15;6:33413. doi: 10.1038/srep33413. Sci Rep. 2016. PMID: 27629591 Free PMC article.
-
Pho4 mediates phosphate acquisition in Candida albicans and is vital for stress resistance and metal homeostasis.Mol Biol Cell. 2016 Sep 1;27(17):2784-801. doi: 10.1091/mbc.E16-05-0266. Epub 2016 Jul 6. Mol Biol Cell. 2016. PMID: 27385340 Free PMC article.
-
Landscape of gene expression variation of natural isolates of Cryptococcus neoformans in response to biologically relevant stresses.Microb Genom. 2020 Jan;6(1):e000319. doi: 10.1099/mgen.0.000319. Microb Genom. 2020. PMID: 31860441 Free PMC article.
-
Stress-Activated Protein Kinases in Human Fungal Pathogens.Front Cell Infect Microbiol. 2019 Jul 17;9:261. doi: 10.3389/fcimb.2019.00261. eCollection 2019. Front Cell Infect Microbiol. 2019. PMID: 31380304 Free PMC article. Review.
-
Stress signaling pathways for the pathogenicity of Cryptococcus.Eukaryot Cell. 2013 Dec;12(12):1564-77. doi: 10.1128/EC.00218-13. Epub 2013 Sep 27. Eukaryot Cell. 2013. PMID: 24078305 Free PMC article. Review.
References
-
- Arino J. Integrative responses to high pH stress in S. cerevisiae. OMICS. 2010;14:517–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
