

Identification of the cglC, cglD, cglE, and cglF genes and their role in cell contact-dependent gliding motility in Myxococcus xanthus
- PMID: 22343295
- PMCID: PMC3318471
- DOI: 10.1128/JB.00055-12
Identification of the cglC, cglD, cglE, and cglF genes and their role in cell contact-dependent gliding motility in Myxococcus xanthus
Abstract
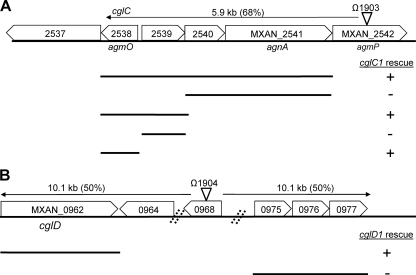
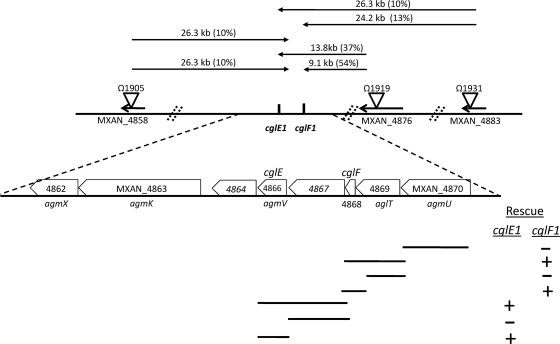
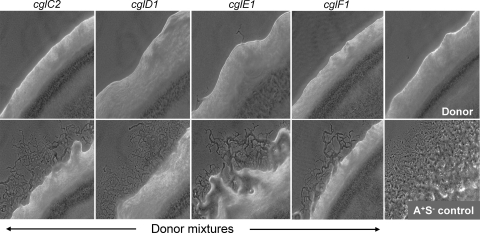
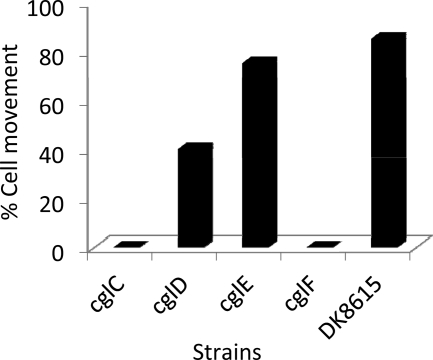
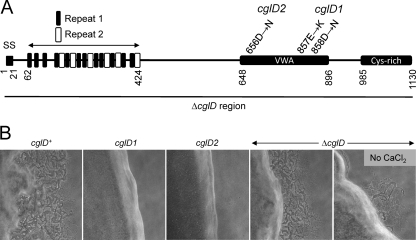
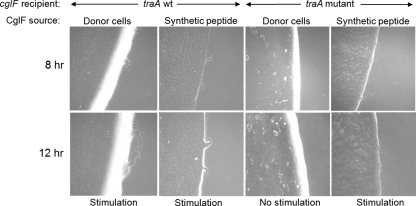
Within Myxococcus xanthus biofilms, cells actively move and exchange their outer membrane (OM) lipoproteins and lipids. Between genetically distinct strains, OM exchange can regulate recipient cell behaviors, including gliding motility and development. Although many different proteins are thought to be exchanged, to date, only two endogenous OM lipoproteins, CglB and Tgl, are known to be transferred. Protein exchange requires the TraAB proteins in recipient and donor cells, where they are hypothesized to facilitate OM fusion for transfer. To better understand the types of proteins exchanged, we identified the genes for the remaining set of cgl gliding motility mutants. These mutants are unique because their motility defect can be transiently restored by physical contact with donor cells that encode the corresponding wild-type protein, a process called stimulation. Similar to CglB and Tgl, the cglC and cglD genes encode type II signal sequences, suggesting that they are also lipoproteins. Surprisingly, the cglE and cglF genes instead encode type I signal sequences, suggesting that nonlipoproteins are also exchanged. Consistent with this idea, the addition of exogenous synthetic CglF protein (71 amino acids) to a cglF mutant rescued its motility defect. In contrast to a live donor cell, stimulation with purified CglF protein occurred independently of TraA. These results also indicate that CglF may localize to the cell surface. The implications of our findings on OM exchange are discussed.
Figures
Similar articles
-
Myxobacterial tools for social interactions.Res Microbiol. 2012 Nov-Dec;163(9-10):579-91. doi: 10.1016/j.resmic.2012.10.022. Epub 2012 Nov 2. Res Microbiol. 2012. PMID: 23123306 Free PMC article. Review.
-
Contact- and Protein Transfer-Dependent Stimulation of Assembly of the Gliding Motility Machinery in Myxococcus xanthus.PLoS Genet. 2015 Jul 1;11(7):e1005341. doi: 10.1371/journal.pgen.1005341. eCollection 2015 Jul. PLoS Genet. 2015. PMID: 26132848 Free PMC article.
-
Cell-to-cell transfer of bacterial outer membrane lipoproteins.Science. 2005 Jul 1;309(5731):125-7. doi: 10.1126/science.1112440. Science. 2005. PMID: 15994555
-
Genetic and molecular analysis of cglB, a gene essential for single-cell gliding in Myxococcus xanthus.J Bacteriol. 1999 Jul;181(14):4381-90. doi: 10.1128/JB.181.14.4381-4390.1999. J Bacteriol. 1999. PMID: 10400597 Free PMC article.
-
Gliding motility in bacteria: insights from studies of Myxococcus xanthus.Microbiol Mol Biol Rev. 1999 Sep;63(3):621-41. doi: 10.1128/MMBR.63.3.621-641.1999. Microbiol Mol Biol Rev. 1999. PMID: 10477310 Free PMC article. Review.
Cited by
-
Self-identity barcodes encoded by six expansive polymorphic toxin families discriminate kin in myxobacteria.Proc Natl Acad Sci U S A. 2019 Dec 3;116(49):24808-24818. doi: 10.1073/pnas.1912556116. Epub 2019 Nov 19. Proc Natl Acad Sci U S A. 2019. PMID: 31744876 Free PMC article.
-
Myxobacteria produce outer membrane-enclosed tubes in unstructured environments.J Bacteriol. 2014 May;196(10):1807-14. doi: 10.1128/JB.00850-13. Epub 2014 Jan 3. J Bacteriol. 2014. PMID: 24391054 Free PMC article.
-
Modular Lipoprotein Toxins Transferred by Outer Membrane Exchange Target Discrete Cell Entry Pathways.mBio. 2021 Oct 26;12(5):e0238821. doi: 10.1128/mBio.02388-21. Epub 2021 Sep 14. mBio. 2021. PMID: 34517761 Free PMC article.
-
Sibling Rivalry in Myxococcus xanthus Is Mediated by Kin Recognition and a Polyploid Prophage.J Bacteriol. 2016 Jan 19;198(6):994-1004. doi: 10.1128/JB.00964-15. J Bacteriol. 2016. PMID: 26787762 Free PMC article.
-
Myxobacterial tools for social interactions.Res Microbiol. 2012 Nov-Dec;163(9-10):579-91. doi: 10.1016/j.resmic.2012.10.022. Epub 2012 Nov 2. Res Microbiol. 2012. PMID: 23123306 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
