

Characterization of Campylobacter jejuni RacRS reveals roles in the heat shock response, motility, and maintenance of cell length homogeneity
- PMID: 22343300
- PMCID: PMC3347078
- DOI: 10.1128/JB.06041-11
Characterization of Campylobacter jejuni RacRS reveals roles in the heat shock response, motility, and maintenance of cell length homogeneity
Abstract
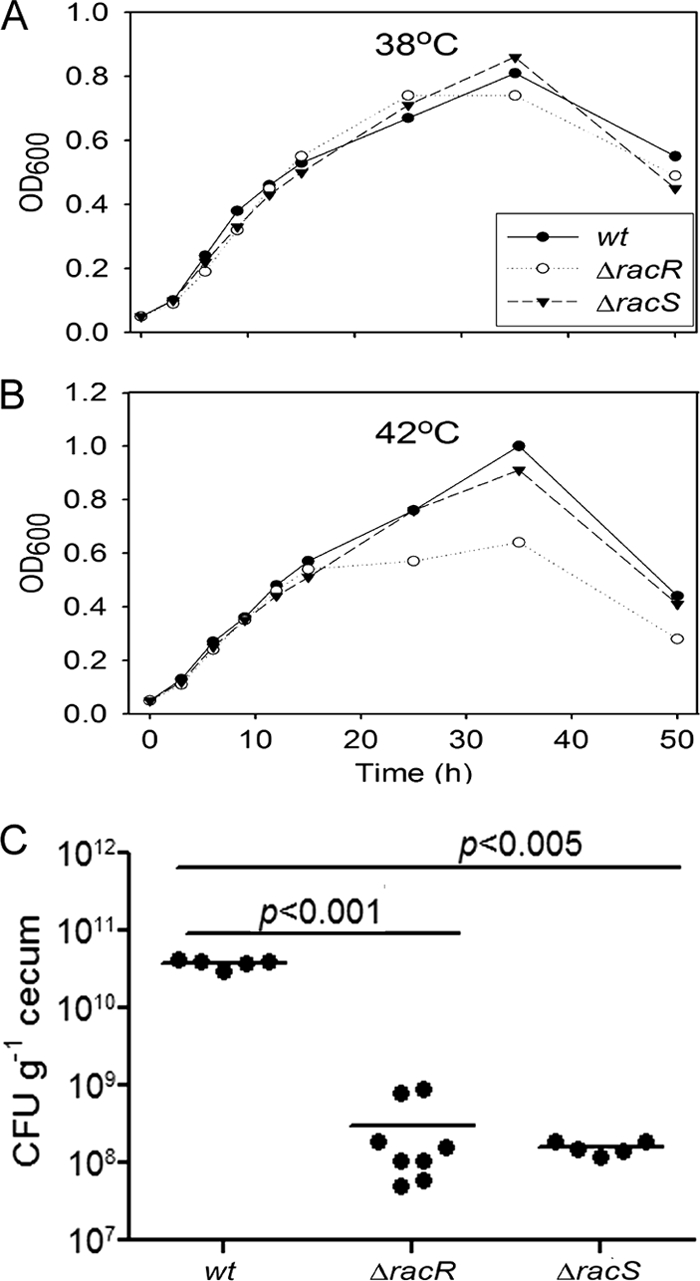
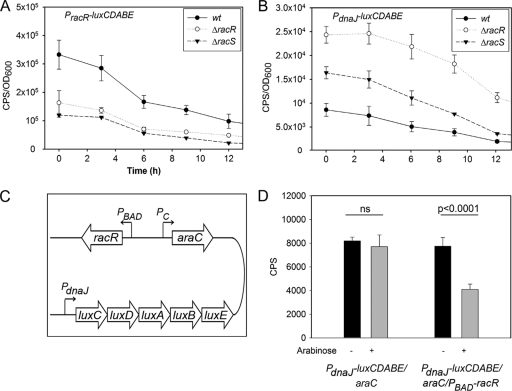
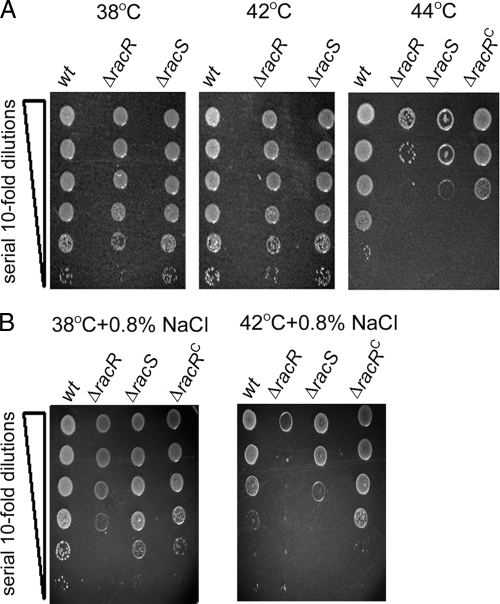
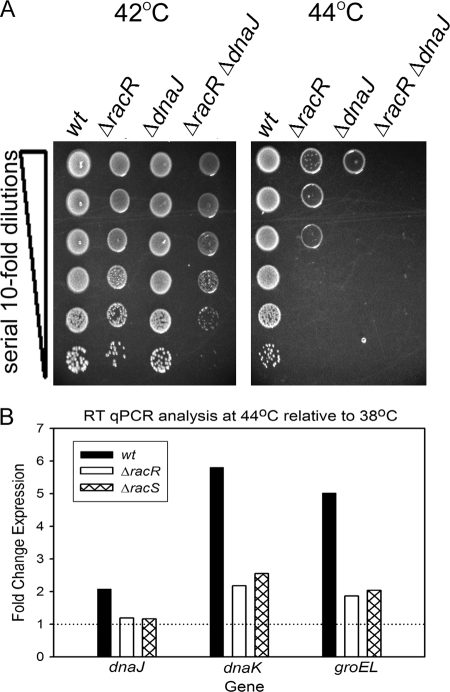
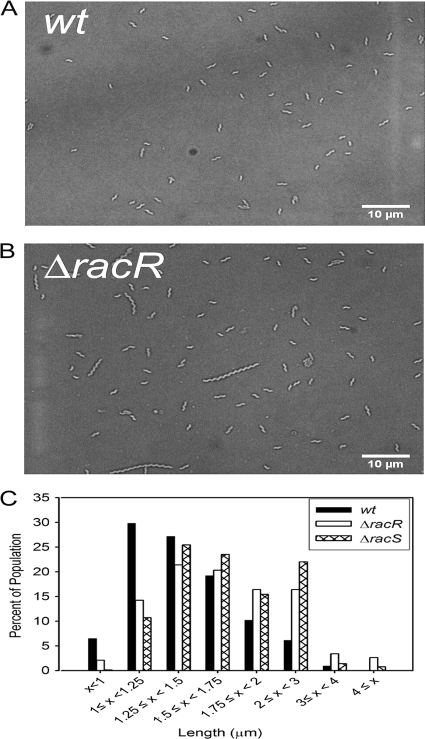
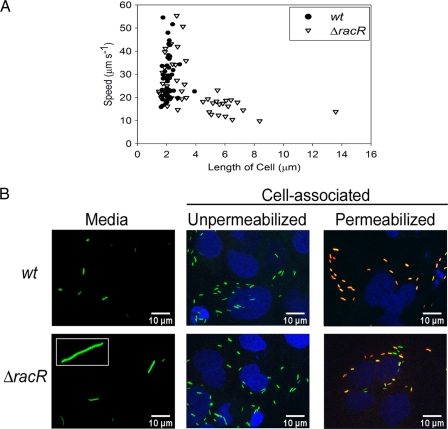
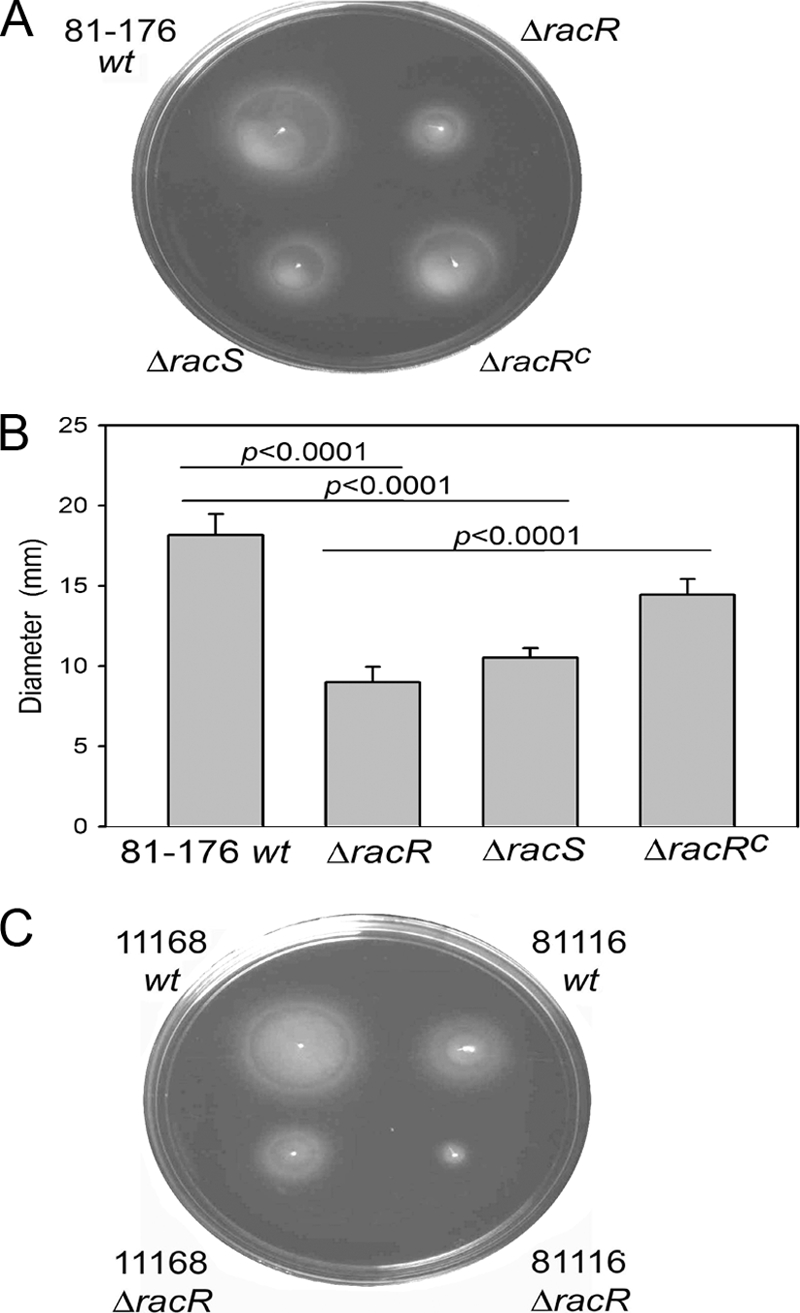
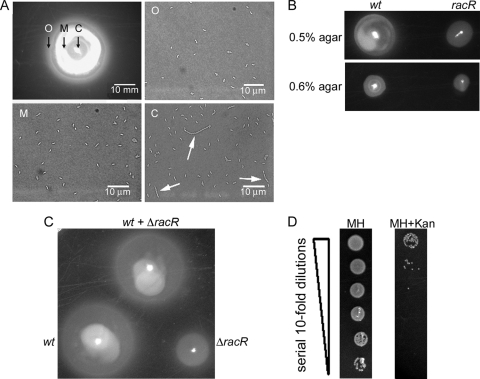
Campylobacter jejuni commensally colonizes the cecum of birds. The RacR (reduced ability to colonize) response regulator was previously shown to be important in avian colonization. To explore the means by which RacR and its cognate sensor kinase RacS may modulate C. jejuni physiology and colonization, ΔracR and ΔracS mutations were constructed in the invasive, virulent strain 81-176, and extensive phenotypic analyses were undertaken. Both the ΔracR and ΔracS mutants exhibited a ~100-fold defect in chick colonization despite no (ΔracS) or minimal (ΔracR) growth defects at 42 °C, the avian body temperature. Each mutant was defective for colony formation at 44°C and in the presence of 0.8% NaCl, both of which are stresses associated with the heat shock response. Promoter-reporter and real-time quantitative PCR (RT-qPCR) analyses revealed that RacR activates racRS and represses dnaJ. Although disregulation of several other heat shock genes was not observed at 38°C, the ΔracR and ΔracS mutants exhibited diminished upregulation of these genes upon a rapid temperature upshift. Furthermore, the ΔracR and ΔracS mutants displayed increased length heterogeneity during exponential growth, with a high proportion of filamented bacteria. Filamented bacteria had reduced swimming speed and were defective for invasion of Caco-2 epithelial cells. Soft-agar studies also revealed that the loss of racR or racS resulted in whole-population motility defects in viscous medium. These findings reveal new roles for RacRS in C. jejuni physiology, each of which is likely important during colonization of the avian host.
Figures
References
-
- Allen KJ, Griffiths MW. 2001. Effect of environmental and chemotactic stimuli on the activity of the Campylobacter jejuni flaA sigma(28) promoter. FEMS Microbiol. Lett. 205:43–48 - PubMed
-
- Allos BM. 1998. Campylobacter jejuni infection as a cause of the Guillain-Barre syndrome. Infect. Dis. Clin. North. Am. 12:173–184 - PubMed
-
- Allos BM. 2001. Campylobacter jejuni infections: update on emerging issues and trends. Clin. Infect. Dis. 32:1201–1206 - PubMed
-
- Andersen MT, et al. 2005. Diverse roles for HspR in Campylobacter jejuni revealed by the proteome, transcriptome and phenotypic characterization of an hspR mutant. Microbiology 151:905–915 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
