

P2X7 receptor-NADPH oxidase axis mediates protein radical formation and Kupffer cell activation in carbon tetrachloride-mediated steatohepatitis in obese mice
- PMID: 22343416
- PMCID: PMC3341527
- DOI: 10.1016/j.freeradbiomed.2012.02.010
P2X7 receptor-NADPH oxidase axis mediates protein radical formation and Kupffer cell activation in carbon tetrachloride-mediated steatohepatitis in obese mice
Abstract
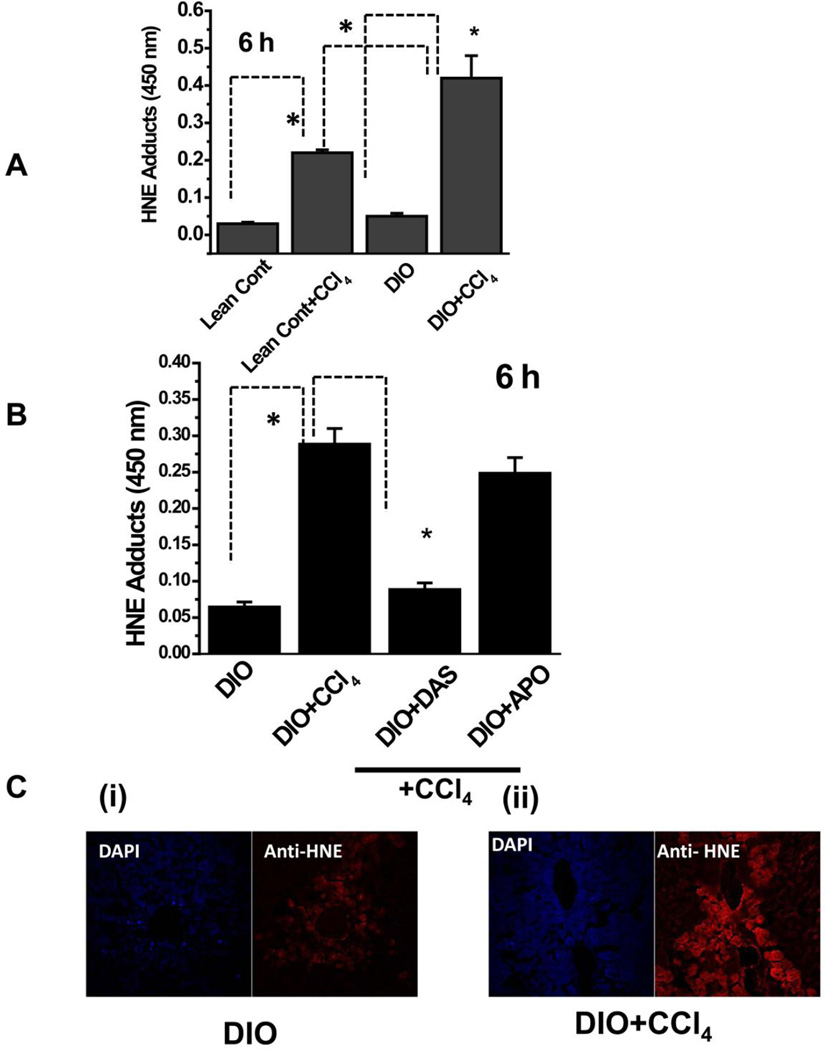
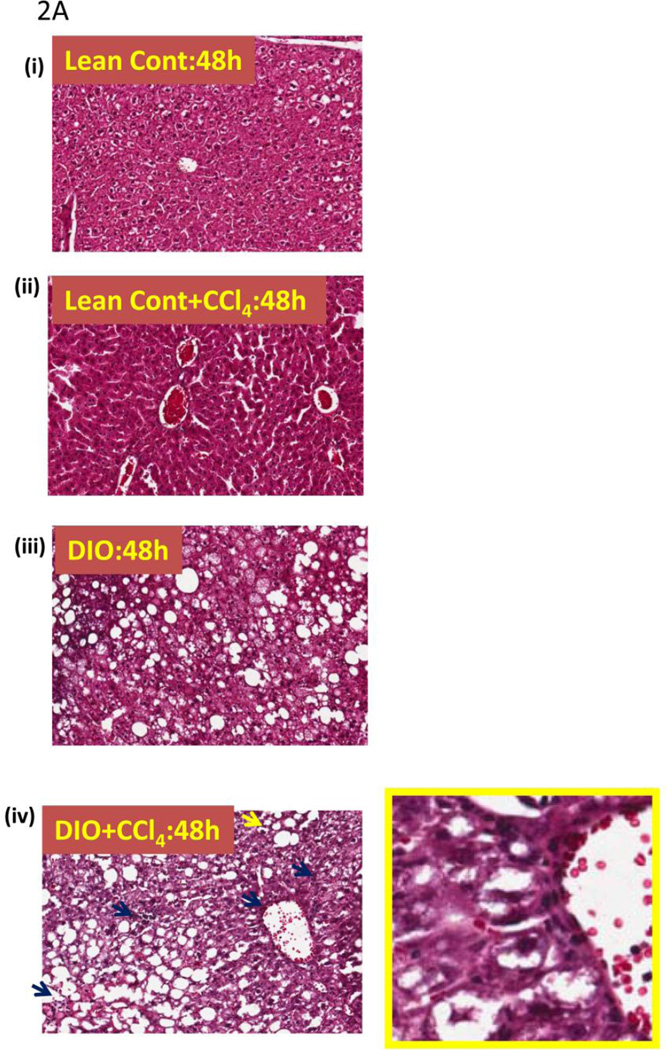
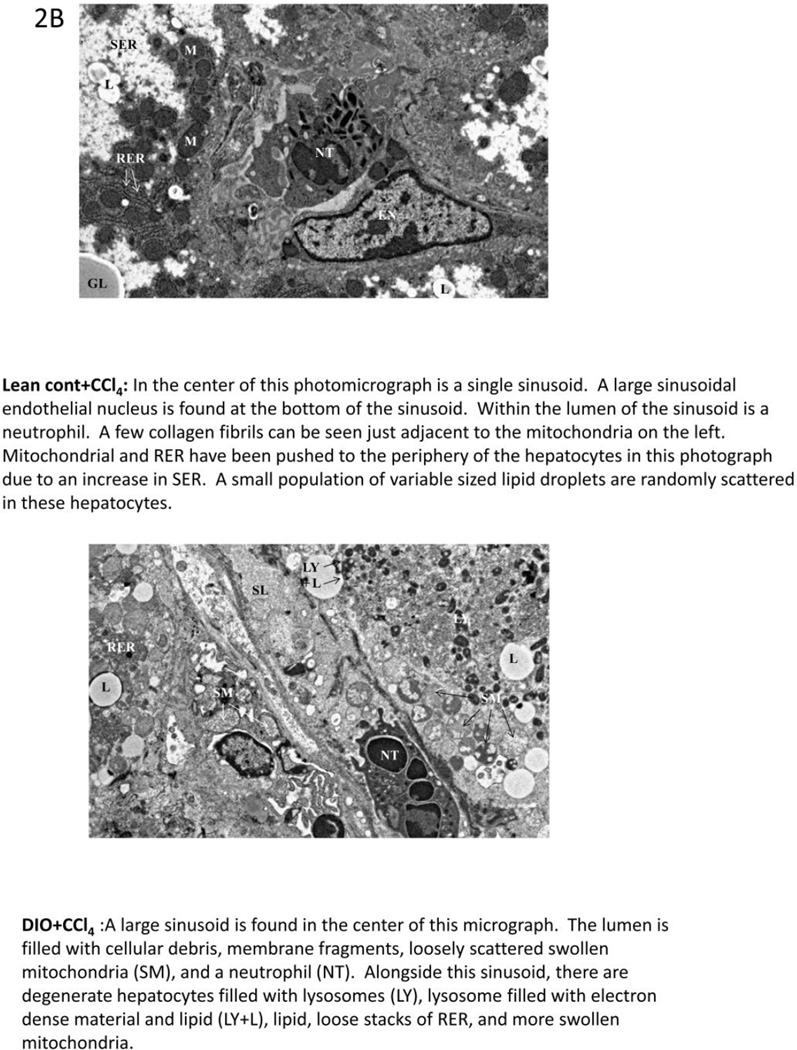
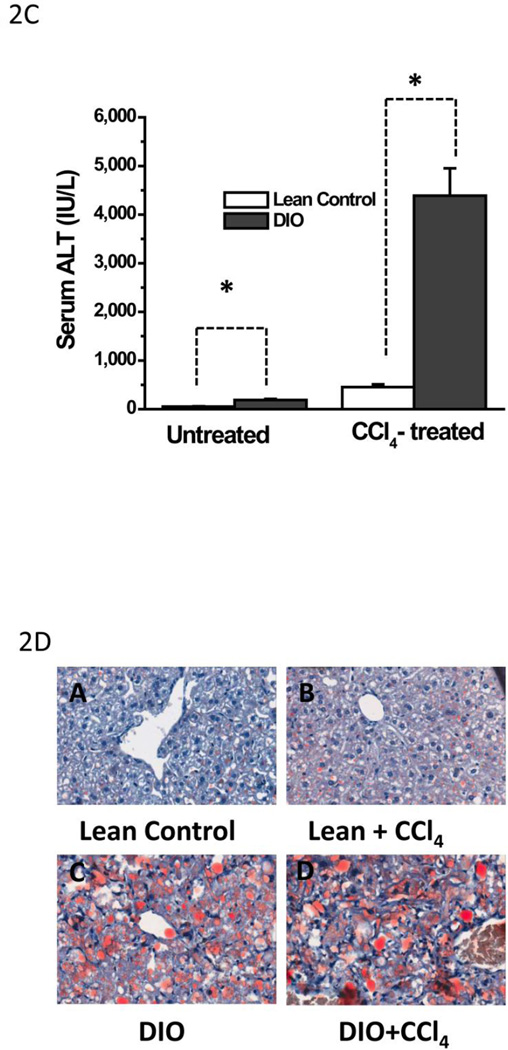
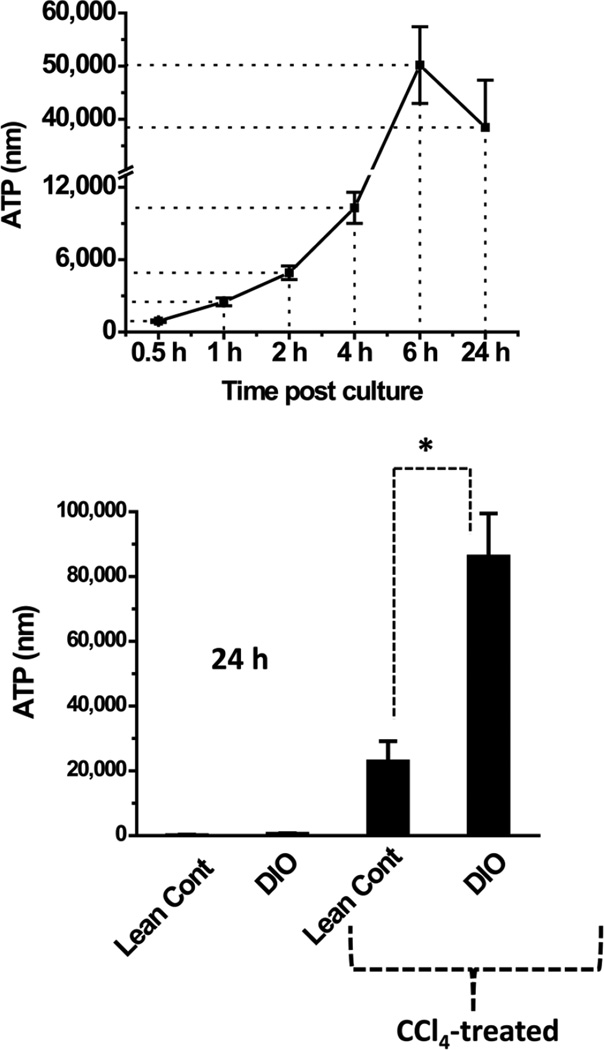
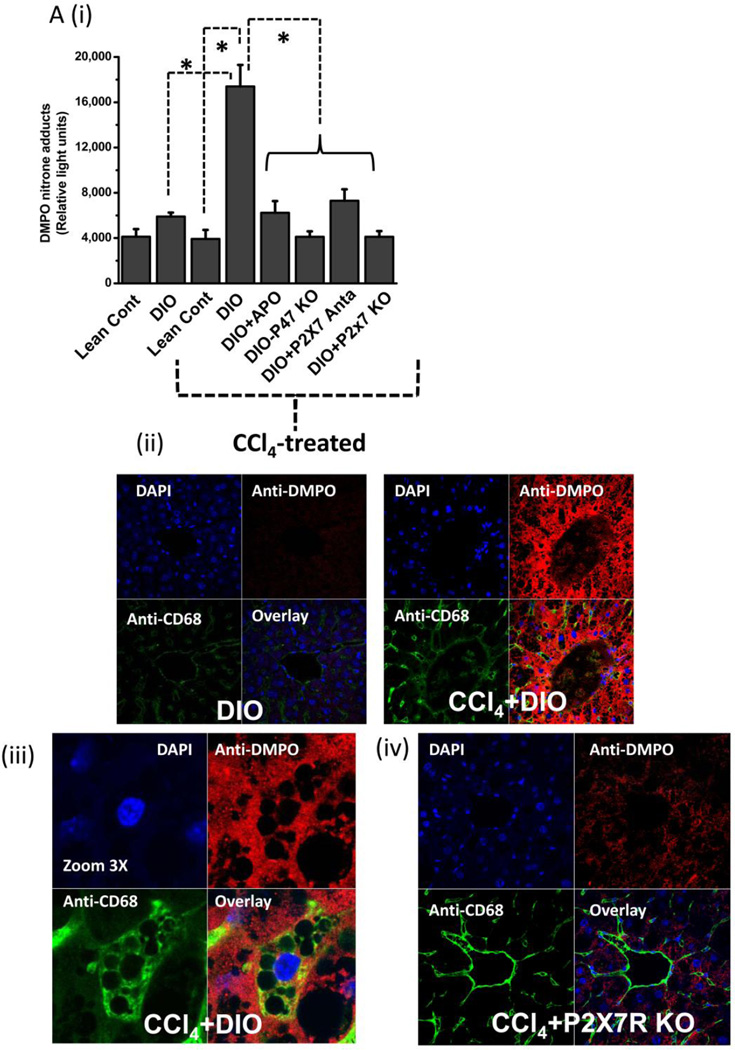
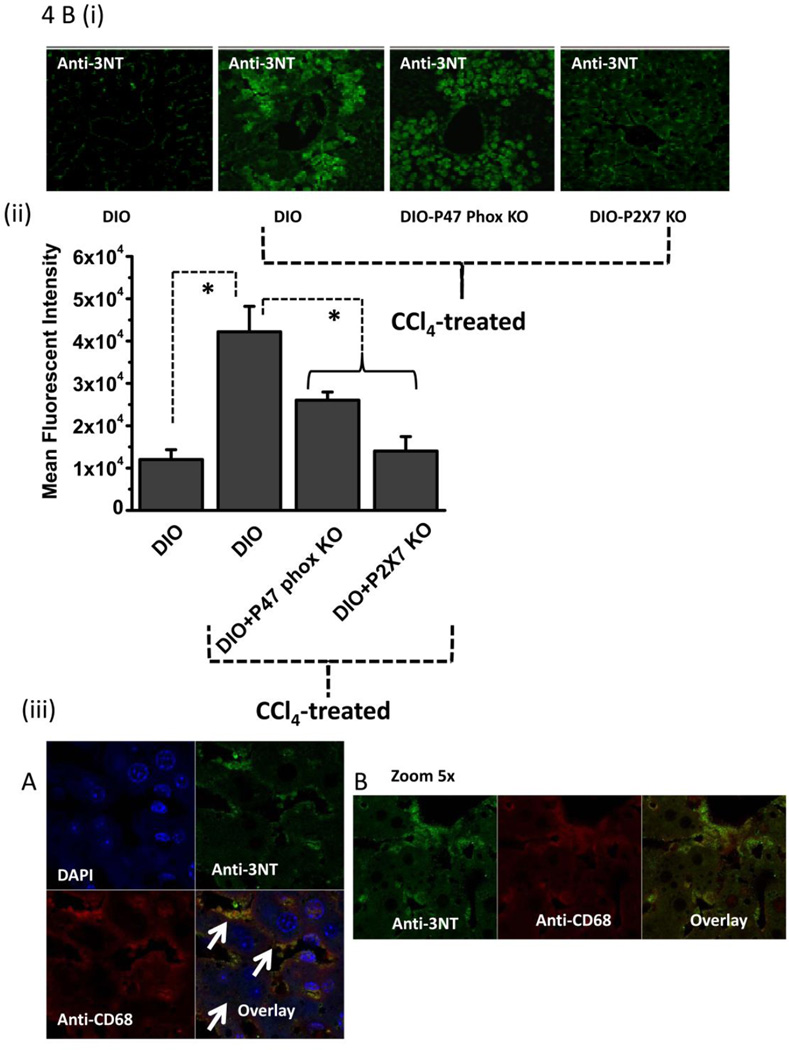
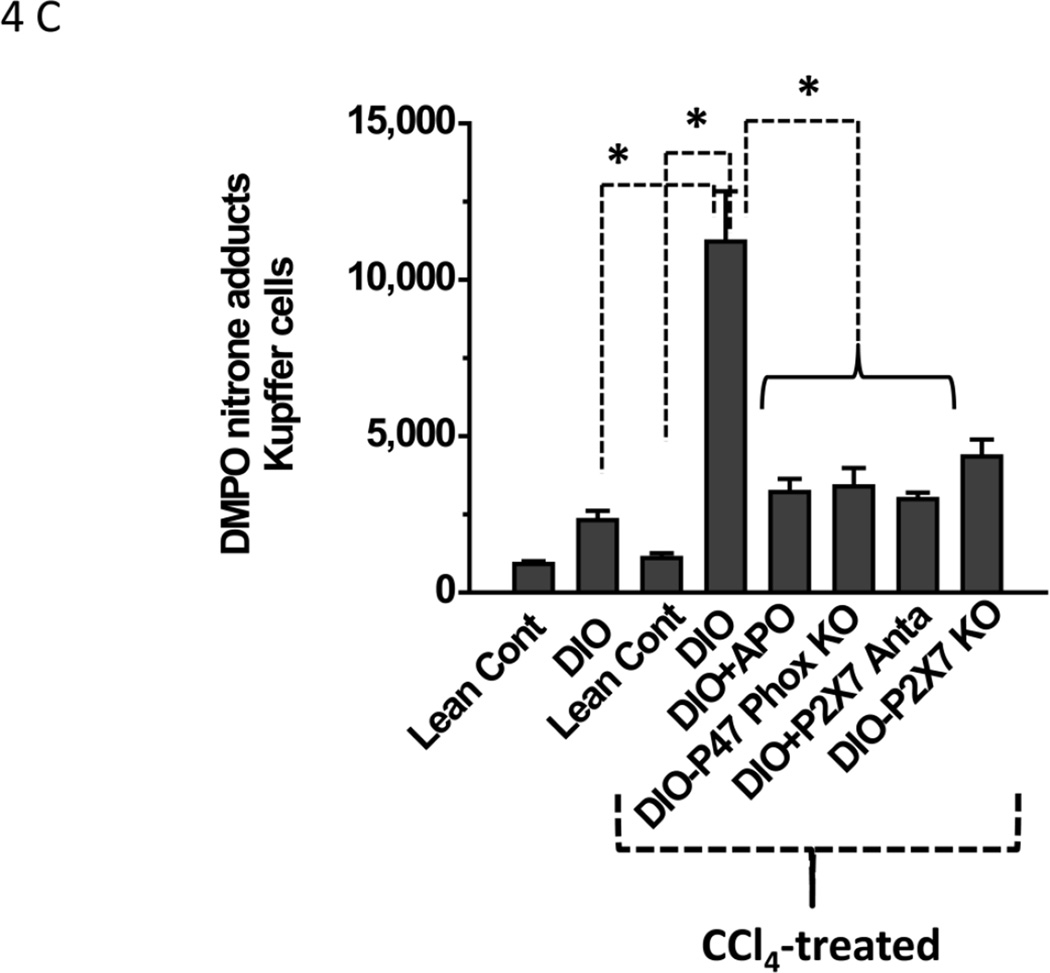
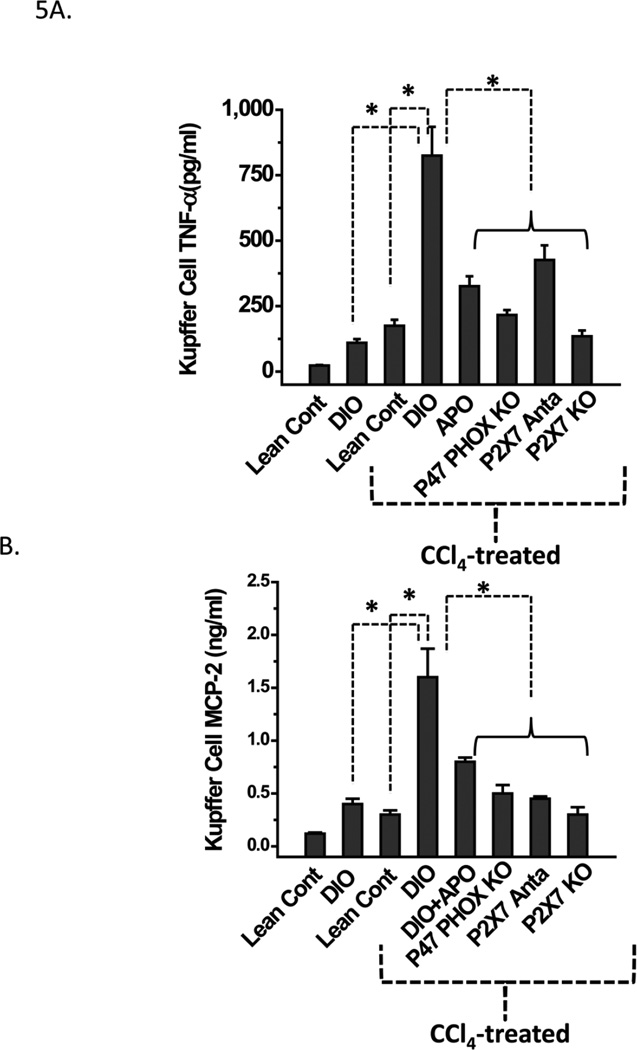
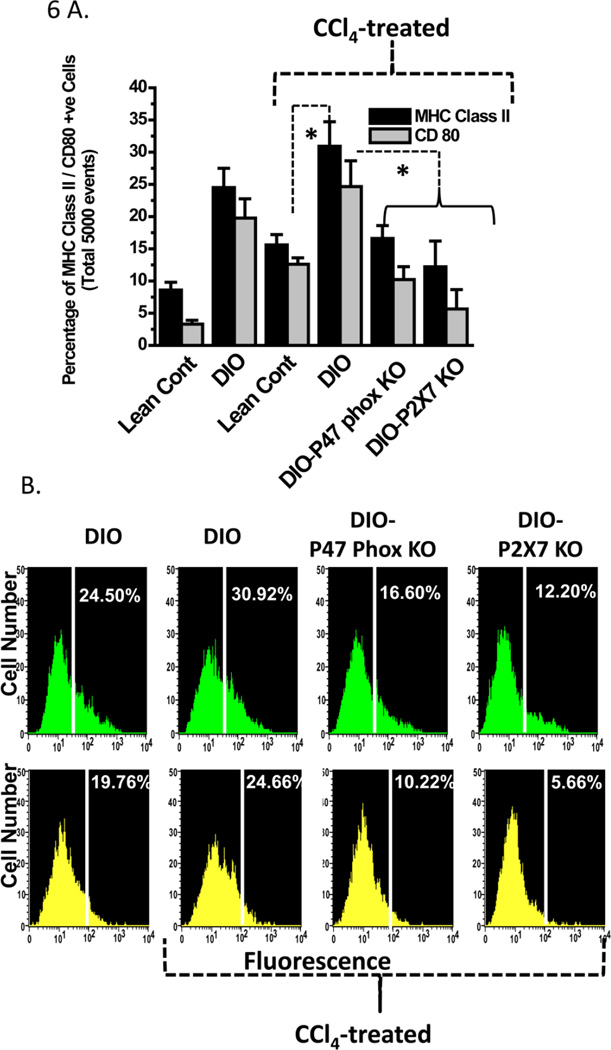
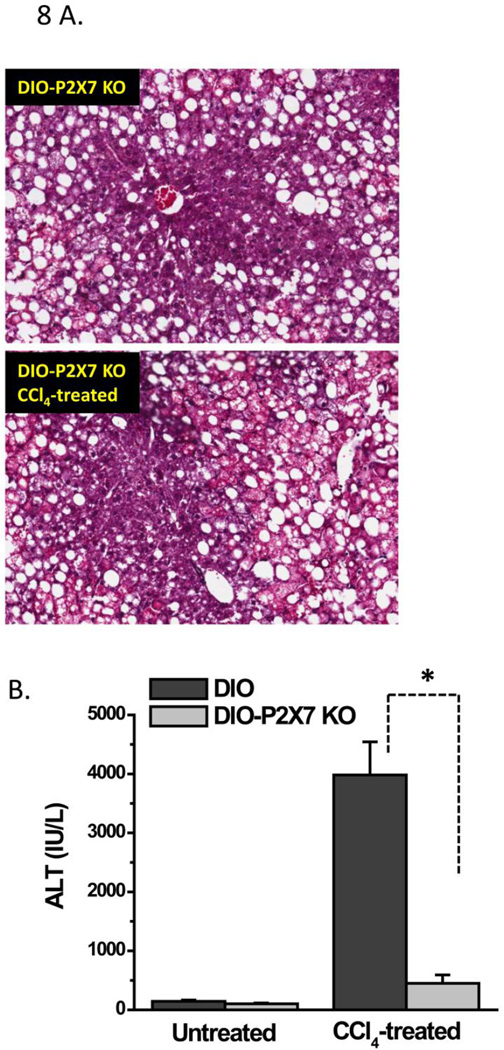
While some studies show that carbon tetrachloride-mediated metabolic oxidative stress exacerbates steatohepatitic-like lesions in obese mice, the redox mechanisms that trigger the innate immune system and accentuate the inflammatory cascade remain unclear. Here we have explored the role of the purinergic receptor P2X7-NADPH oxidase axis as a primary event in recognizing the heightened release of extracellular ATP from CCl(4)-treated hepatocytes and generating redox-mediated Kupffer cell activation in obese mice. We found that an underlying condition of obesity led to the formation of protein radicals and posttranslational nitration, primarily in Kupffer cells, at 24h post-CCl(4) administration. The free radical-mediated oxidation of cellular macromolecules, which was NADPH oxidase and P2X7 receptor-dependent, correlated well with the release of TNF-α and MCP-2 from Kupffer cells. The Kupffer cells in CCl(4)-treated mice exhibited increased expression of MHC Class II proteins and showed an activated phenotype. Increased expression of MHC Class II was inhibited by the NADPH oxidase inhibitor apocynin , P2X7 receptor antagonist A438709 hydrochloride, and genetic deletions of the NADPH oxidase p47 phox subunit or the P2X7 receptor. The P2X7 receptor acted upstream of NADPH oxidase activation by up-regulating the expression of the p47 phox subunit and p47 phox binding to the membrane subunit, gp91 phox. We conclude that the P2X7 receptor is a primary mediator of oxidative stress-induced exacerbation of inflammatory liver injury in obese mice via NADPH oxidase-dependent mechanisms.
Published by Elsevier Inc.
Figures

References
-
- Farrell GC, Larter CZ. Nonalcoholic fatty liver disease: from steatosis to cirrhosis. Hepatology. 2006. 2006 Feb;43(2 Suppl 1):S99–S112. - PubMed
-
- Diehl AM. Hepatic complications of obesity. Gastroenterol Clin North Am. 2010. 2010 Mar;39(1):57–68. - PubMed
-
- Pessayre D. Role of mitochondria in non-alcoholic fatty liver disease. J Gastroenterol Hepatol. 2007;22(Suppl 1):S20–S27. - PubMed
-
- Fromenty B, Fisch C, Labbe G, Degott C, Deschamps D, Berson A, Letteron P, Pessayre D. Amiodarone inhibits the mitochondrial beta-oxidation of fatty acids and produces microvesicular steatosis of the liver in mice. J Pharmacol Exp Ther. 1990;255(3):1371–1376. - PubMed
-
- Fromenty B, Fisch C, Berson A, Letteron P, Larrey D, Pessayre D. Dual effect of amiodarone on mitochondrial respiration. Initial protonophoric uncoupling effect followed by inhibition of the respiratory chain at the levels of complex I and complex II. J Pharmacol Exp Ther. 1990;255(3):1377–1384. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
