

The role of Drosophila Piezo in mechanical nociception
- PMID: 22343891
- PMCID: PMC3297676
- DOI: 10.1038/nature10801
The role of Drosophila Piezo in mechanical nociception
Abstract
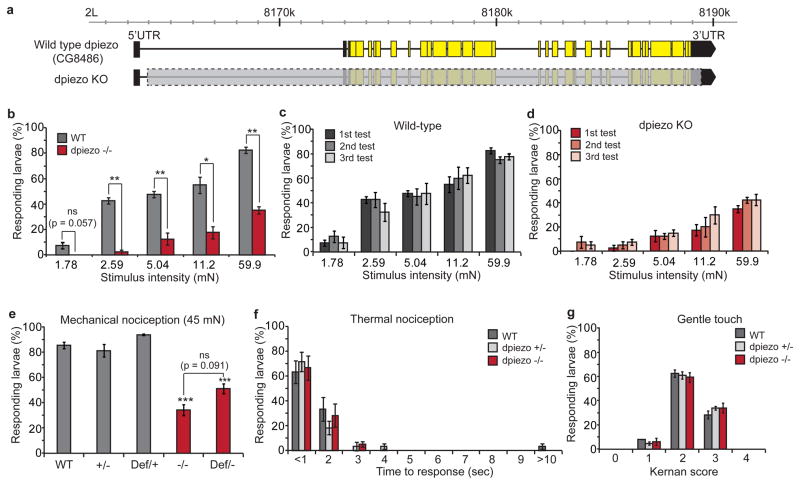
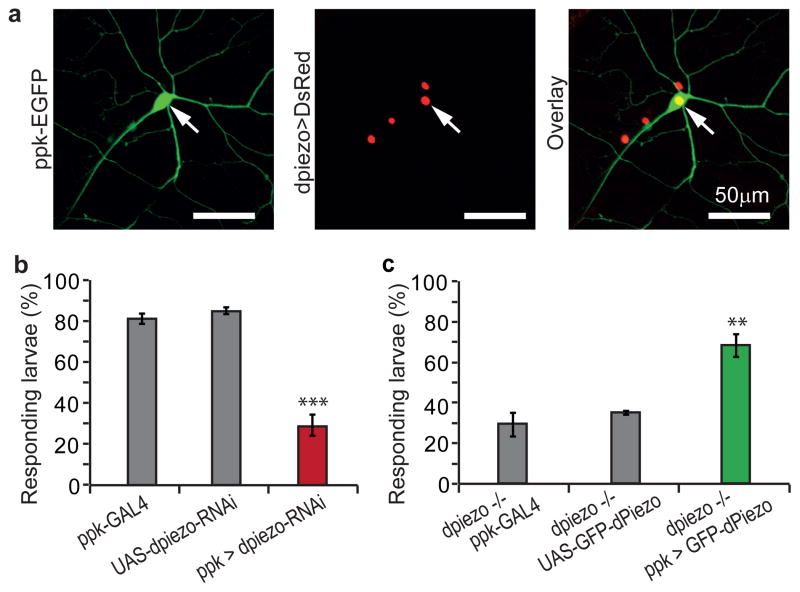
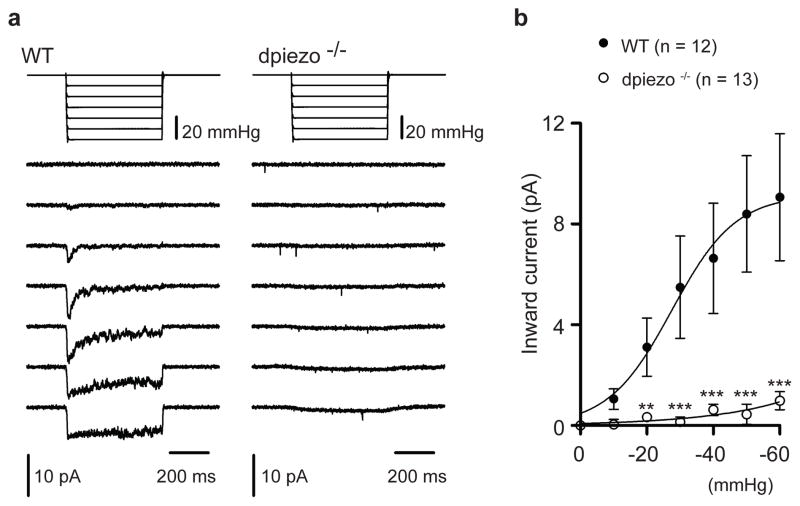
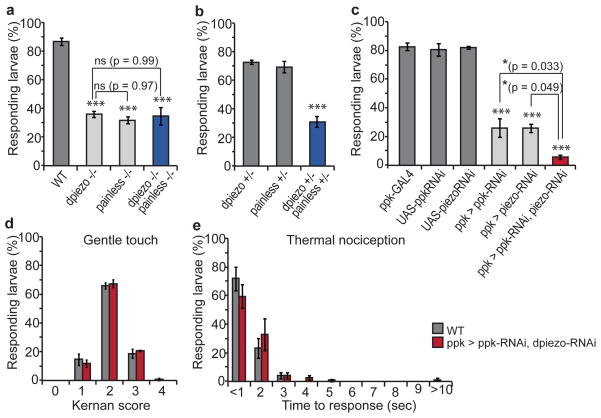
Transduction of mechanical stimuli by receptor cells is essential for senses such as hearing, touch and pain. Ion channels have a role in neuronal mechanotransduction in invertebrates; however, functional conservation of these ion channels in mammalian mechanotransduction is not observed. For example, no mechanoreceptor potential C (NOMPC), a member of transient receptor potential (TRP) ion channel family, acts as a mechanotransducer in Drosophila melanogaster and Caenorhabditis elegans; however, it has no orthologues in mammals. Degenerin/epithelial sodium channel (DEG/ENaC) family members are mechanotransducers in C. elegans and potentially in D. melanogaster; however, a direct role of its mammalian homologues in sensing mechanical force has not been shown. Recently, Piezo1 (also known as Fam38a) and Piezo2 (also known as Fam38b) were identified as components of mechanically activated channels in mammals. The Piezo family are evolutionarily conserved transmembrane proteins. It is unknown whether they function in mechanical sensing in vivo and, if they do, which mechanosensory modalities they mediate. Here we study the physiological role of the single Piezo member in D. melanogaster (Dmpiezo; also known as CG8486). Dmpiezo expression in human cells induces mechanically activated currents, similar to its mammalian counterparts. Behavioural responses to noxious mechanical stimuli were severely reduced in Dmpiezo knockout larvae, whereas responses to another noxious stimulus or touch were not affected. Knocking down Dmpiezo in sensory neurons that mediate nociception and express the DEG/ENaC ion channel pickpocket (ppk) was sufficient to impair responses to noxious mechanical stimuli. Furthermore, expression of Dmpiezo in these same neurons rescued the phenotype of the constitutive Dmpiezo knockout larvae. Accordingly, electrophysiological recordings from ppk-positive neurons revealed a Dmpiezo-dependent, mechanically activated current. Finally, we found that Dmpiezo and ppk function in parallel pathways in ppk-positive cells, and that mechanical nociception is abolished in the absence of both channels. These data demonstrate the physiological relevance of the Piezo family in mechanotransduction in vivo, supporting a role of Piezo proteins in mechanosensory nociception.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
Cell biology: The sensation of stretch.Nature. 2012 Mar 7;483(7388):163-4. doi: 10.1038/483163a. Nature. 2012. PMID: 22398551 Free PMC article.
References
-
- Chalfie M. Neurosensory mechanotransduction. Nat Rev Mol Cell Biol. 2009;10:44–52. - PubMed
-
- Delmas P, Hao J, Rodat-Despoix L. Molecular mechanisms of mechanotransduction in mammalian sensory neurons. Nat Rev Neurosci. 2011;12:139–153. - PubMed
-
- Walker RG, Willingham AT, Zuker CS. A Drosophila Mechanosensory Transduction Channel. Science. 2000;287:2229–2234. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
