

The hitchhiker's guide to Xenopus genetics
- PMID: 22344745
- PMCID: PMC3312310
- DOI: 10.1002/dvg.22007
The hitchhiker's guide to Xenopus genetics
Abstract
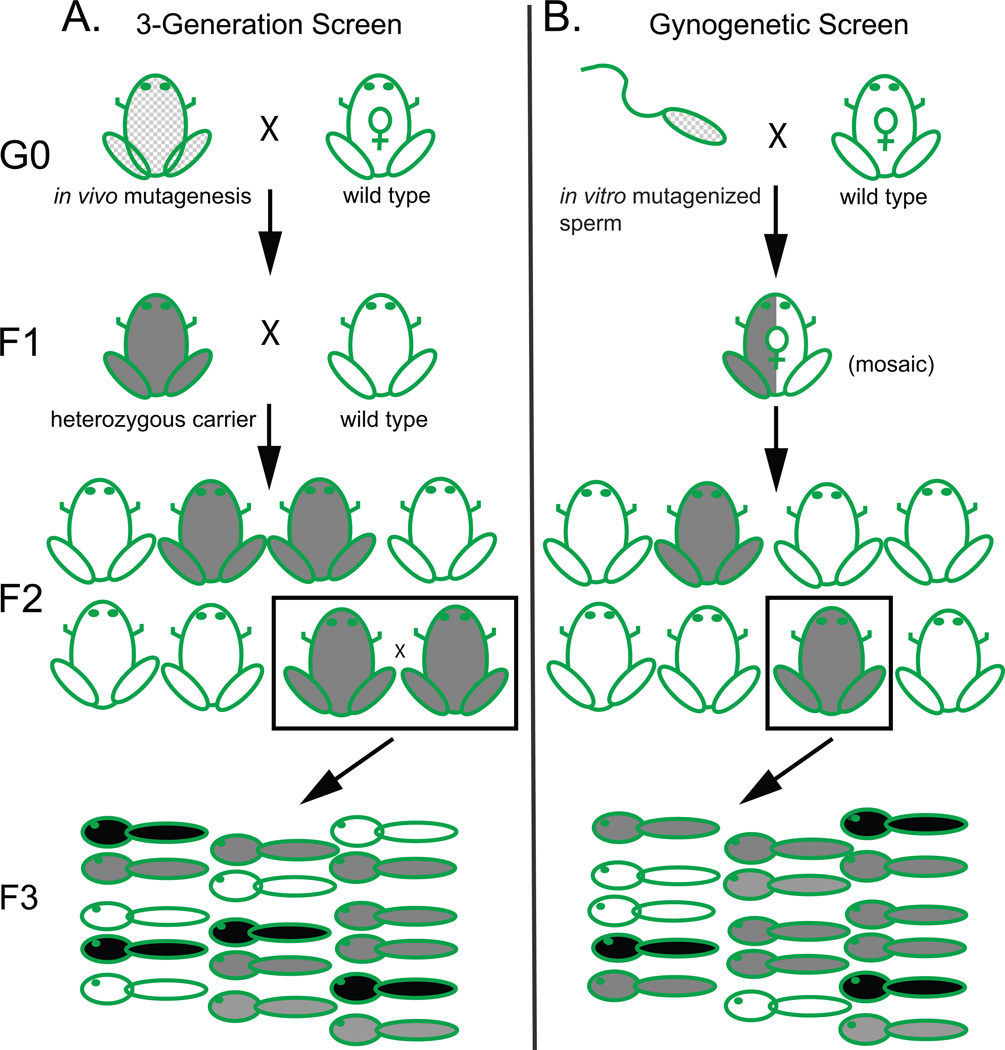
A decade after the human genome sequence, most vertebrate gene functions remain poorly understood, limiting benefits to human health from rapidly advancing genomic technologies. Systematic in vivo functional analysis is ideally suited to the experimentally accessible Xenopus embryo, which combines embryological accessibility with a broad range of transgenic, biochemical, and gain-of-function assays. The diploid X. tropicalis adds loss-of-function genetics and enhanced genomics to this repertoire. In the last decade, diverse phenotypes have been recovered from genetic screens, mutations have been cloned, and reverse genetics in the form of TILLING and targeted gene editing have been established. Simple haploid genetics and gynogenesis and the very large number of embryos produced streamline screening and mapping. Improved genomic resources and the revolution in high-throughput sequencing are transforming mutation cloning and reverse genetic approaches. The combination of loss-of-function mutant backgrounds with the diverse array of conventional Xenopus assays offers a uniquely flexible platform for analysis of gene function in vertebrate development.
Copyright © 2012 Wiley Periodicals, Inc.
Figures



References
-
- Adams D. Life, The Universe and Everything. 1982
-
- Amaya E, Musci TJ, Kirschner MW. Expression of a dominant negative mutant of the FGF receptor disrupts mesoderm formation in Xenopus embryos. Cell. 1991;66:257–270. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
