

JUNGBRUNNEN1, a reactive oxygen species-responsive NAC transcription factor, regulates longevity in Arabidopsis
- PMID: 22345491
- PMCID: PMC3315228
- DOI: 10.1105/tpc.111.090894
JUNGBRUNNEN1, a reactive oxygen species-responsive NAC transcription factor, regulates longevity in Arabidopsis
Abstract
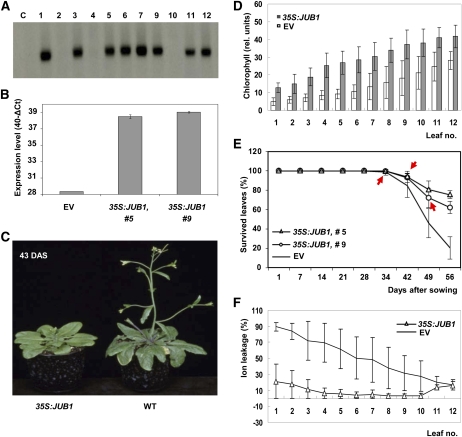
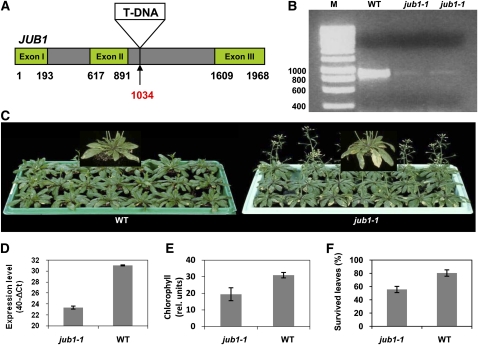
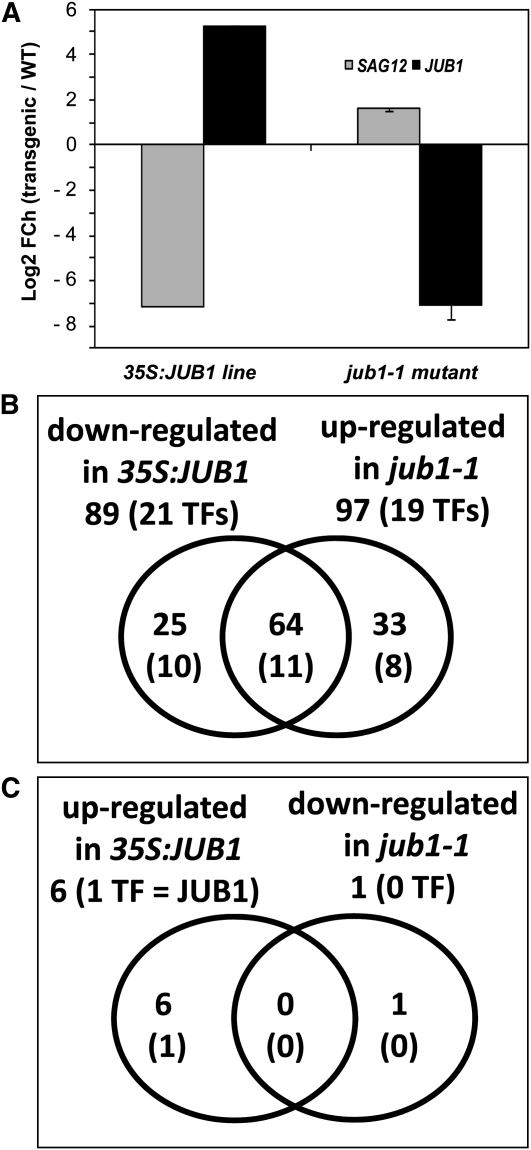
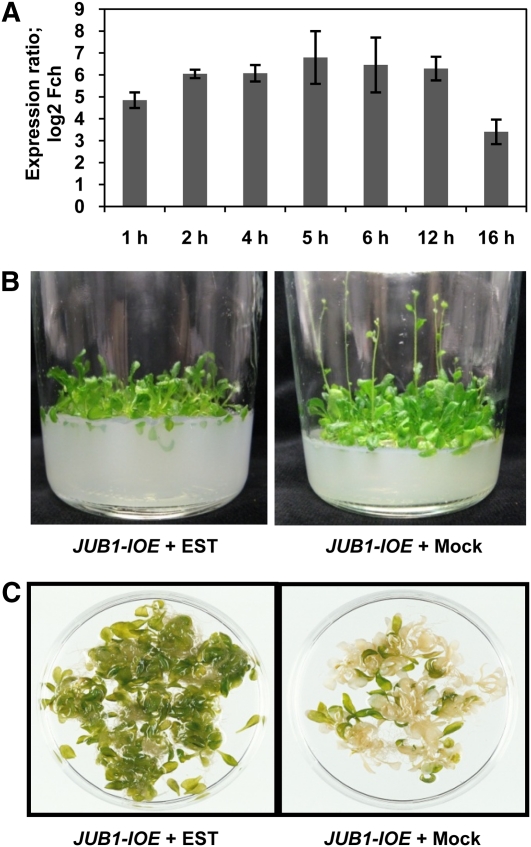
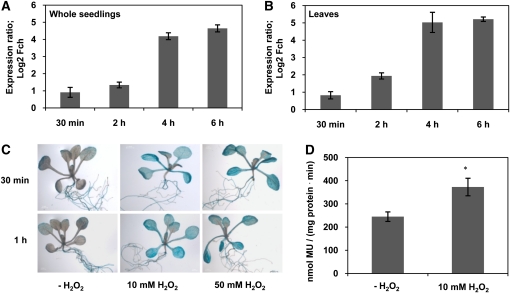
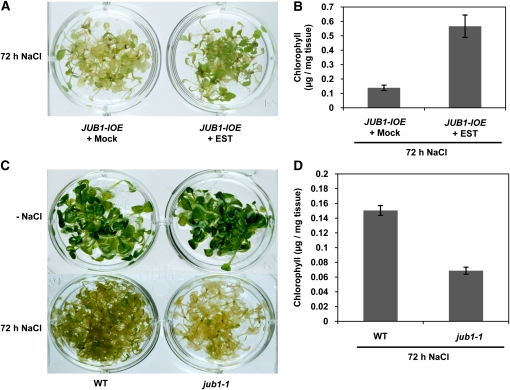
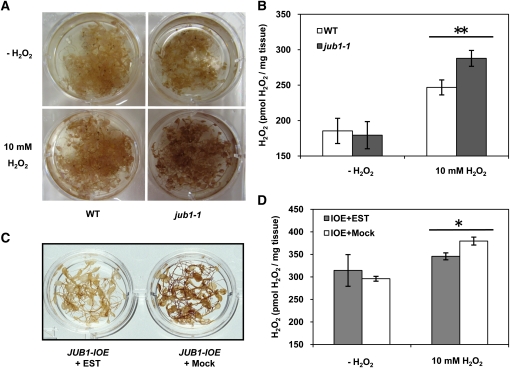
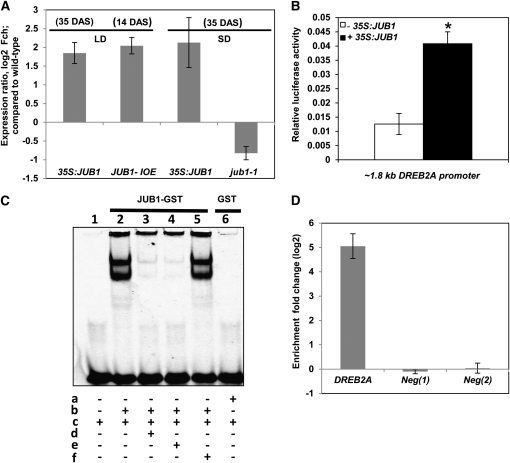
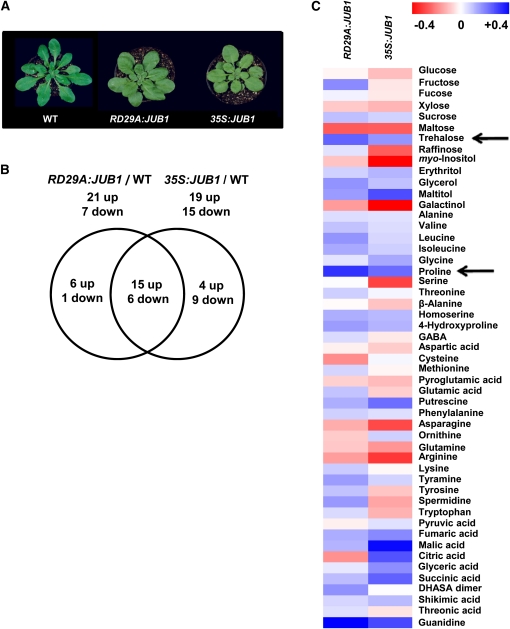
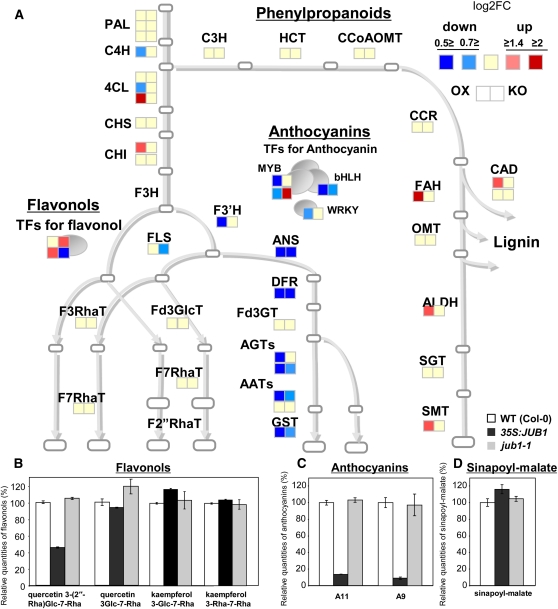
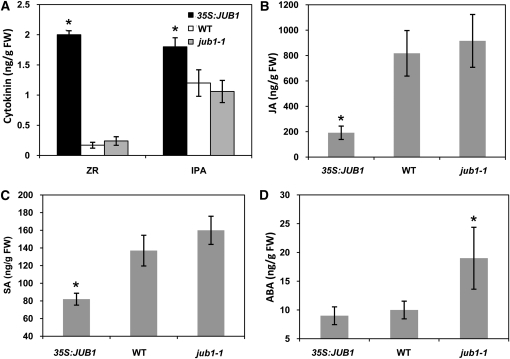
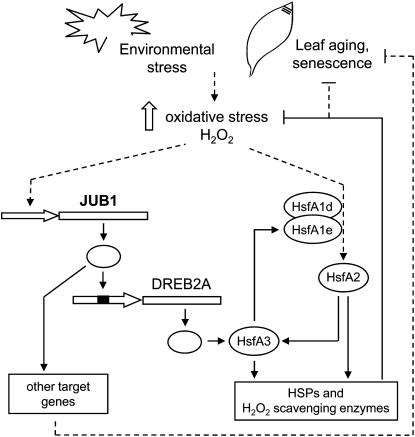
The transition from juvenility through maturation to senescence is a complex process that involves the regulation of longevity. Here, we identify JUNGBRUNNEN1 (JUB1), a hydrogen peroxide (H(2)O(2))-induced NAC transcription factor, as a central longevity regulator in Arabidopsis thaliana. JUB1 overexpression strongly delays senescence, dampens intracellular H(2)O(2) levels, and enhances tolerance to various abiotic stresses, whereas in jub1-1 knockdown plants, precocious senescence and lowered abiotic stress tolerance are observed. A JUB1 binding site containing a RRYGCCGT core sequence is present in the promoter of DREB2A, which plays an important role in abiotic stress responses. JUB1 transactivates DREB2A expression in mesophyll cell protoplasts and transgenic plants and binds directly to the DREB2A promoter. Transcriptome profiling of JUB1 overexpressors revealed elevated expression of several reactive oxygen species-responsive genes, including heat shock protein and glutathione S-transferase genes, whose expression is further induced by H(2)O(2) treatment. Metabolite profiling identified elevated Pro and trehalose levels in JUB1 overexpressors, in accordance with their enhanced abiotic stress tolerance. We suggest that JUB1 constitutes a central regulator of a finely tuned control system that modulates cellular H(2)O(2) level and primes the plants for upcoming stress through a gene regulatory network that involves DREB2A.
Figures


References
-
- Balazadeh S., Parlitz S., Mueller-Roeber B., Meyer R.C. (2008a). Natural developmental variations in leaf and plant senescence in Arabidopsis thaliana. Plant Biol. (Stuttg.) 10(suppl. 1): 136–147 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
