

Human cytomegalovirus UL40 signal peptide regulates cell surface expression of the NK cell ligands HLA-E and gpUL18
- PMID: 22345649
- PMCID: PMC3303119
- DOI: 10.4049/jimmunol.1102068
Human cytomegalovirus UL40 signal peptide regulates cell surface expression of the NK cell ligands HLA-E and gpUL18
Abstract
Human CMV (HCMV)-encoded NK cell-evasion functions include an MHC class I homolog (UL18) with high affinity for the leukocyte inhibitory receptor-1 (CD85j, ILT2, or LILRB1) and a signal peptide (SP(UL40)) that acts by upregulating cell surface expression of HLA-E. Detailed characterization of SP(UL40) revealed that the N-terminal 14 aa residues bestowed TAP-independent upregulation of HLA-E, whereas C region sequences delayed processing of SP(UL40) by a signal peptide peptidase-type intramembrane protease. Most significantly, the consensus HLA-E-binding epitope within SP(UL40) was shown to promote cell surface expression of both HLA-E and gpUL18. UL40 was found to possess two transcription start sites, with utilization of the downstream site resulting in translation being initiated within the HLA-E-binding epitope (P2). Remarkably, this truncated SP(UL40) was functional and retained the capacity to upregulate gpUL18 but not HLA-E. Thus, our findings identify an elegant mechanism by which an HCMV signal peptide differentially regulates two distinct NK cell-evasion pathways. Moreover, we describe a natural SP(UL40) mutant that provides a clear example of an HCMV clinical virus with a defect in an NK cell-evasion function and exemplifies issues that confront the virus when adapting to immunogenetic diversity in the host.
Figures
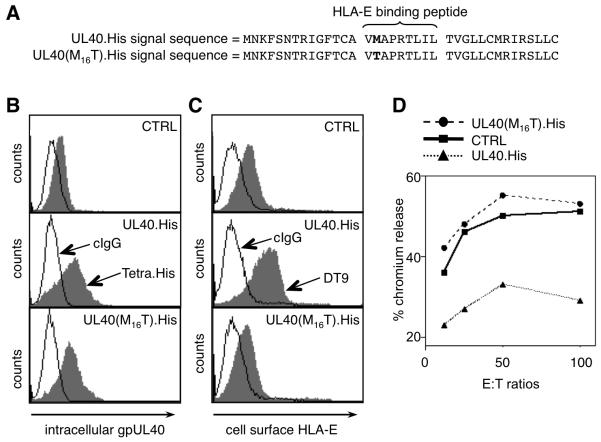

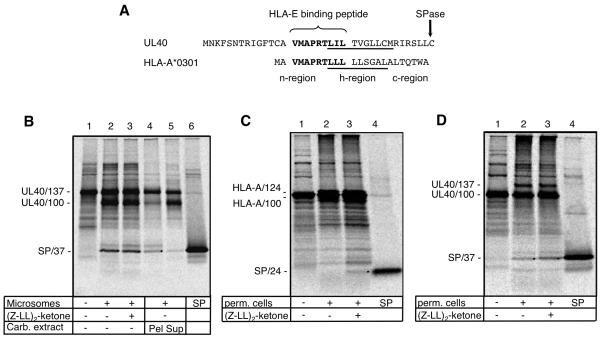
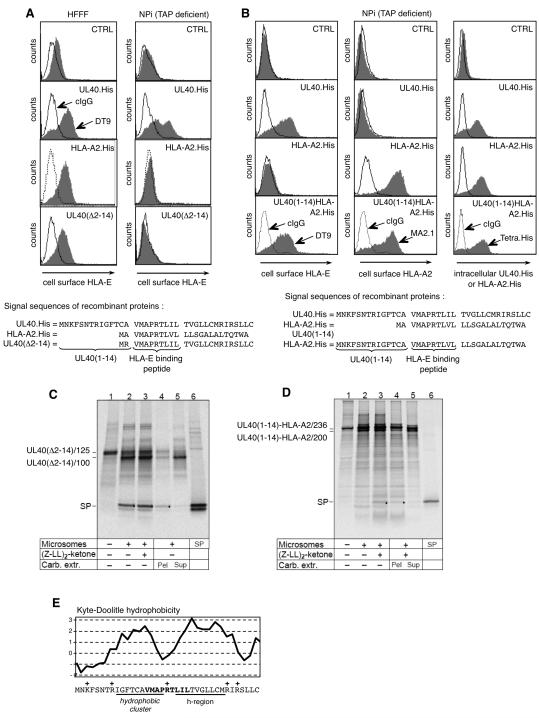

 ), Endo H-sensitive gpUL18 glycoform before (
), Endo H-sensitive gpUL18 glycoform before ( ) and after (
) and after ( ) digestion, PNGase-digested gpUL18 forms (
) digestion, PNGase-digested gpUL18 forms ( ) are indicated. (H) Following cell surface biotinylation and immunoprecipitation, gpUL18, HLA-E, HLA-HC and CD155 cell surface expression were compared on western blot in cells infected with AD169ΔUL16, AD169ΔUL18 or AD169ΔUL40 (15 PFU/cell) for 96h.
) are indicated. (H) Following cell surface biotinylation and immunoprecipitation, gpUL18, HLA-E, HLA-HC and CD155 cell surface expression were compared on western blot in cells infected with AD169ΔUL16, AD169ΔUL18 or AD169ΔUL40 (15 PFU/cell) for 96h.


References
-
- McSharry BP, Burgert HG, Owen DP, Stanton RJ, Prod’homme V, Sester M, Koebernick K, Groh V, Spies T, Cox S, Little AM, Wang EC, Tomasec P, Wilkinson GW. Adenovirus E3/19K promotes evasion of NK cell recognition by intracellular sequestration of the NKG2D ligands major histocompatibility complex class I chain-related proteins A and B. J Virol. 2008;82:4585–4594. - PMC - PubMed
-
- Mocarski ES., Jr. Immunomodulation by cytomegaloviruses: manipulative strategies beyond evasion. Trends Microbiol. 2002;10:332–339. - PubMed
-
- Yokoyama WM, Kim S, French AR. The dynamic life of natural killer cells. Annu Rev Immunol. 2004;22:405–429. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- G0901119(91900)/MRC_/Medical Research Council/United Kingdom
- G1000236/MRC_/Medical Research Council/United Kingdom
- WT090323MA/WT_/Wellcome Trust/United Kingdom
- 090323/WT_/Wellcome Trust/United Kingdom
- MC_U130184136/MRC_/Medical Research Council/United Kingdom
- G0700142(81365)/MRC_/Medical Research Council/United Kingdom
- G0901119/MRC_/Medical Research Council/United Kingdom
- BBF0098361/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G0700142/MRC_/Medical Research Council/United Kingdom
- BB/F009836/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G1000236(94863)/MRC_/Medical Research Council/United Kingdom
- G0801822/MRC_/Medical Research Council/United Kingdom
- BBS/B/02525/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
