

Genome-wide analysis of the world's sheep breeds reveals high levels of historic mixture and strong recent selection
- PMID: 22346734
- PMCID: PMC3274507
- DOI: 10.1371/journal.pbio.1001258
Genome-wide analysis of the world's sheep breeds reveals high levels of historic mixture and strong recent selection
Abstract
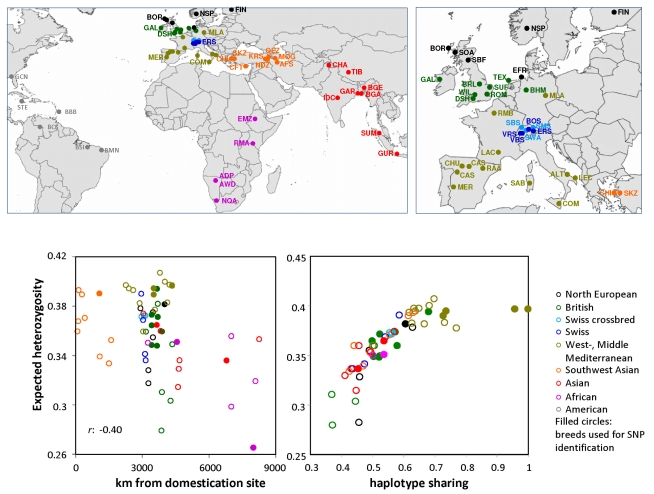
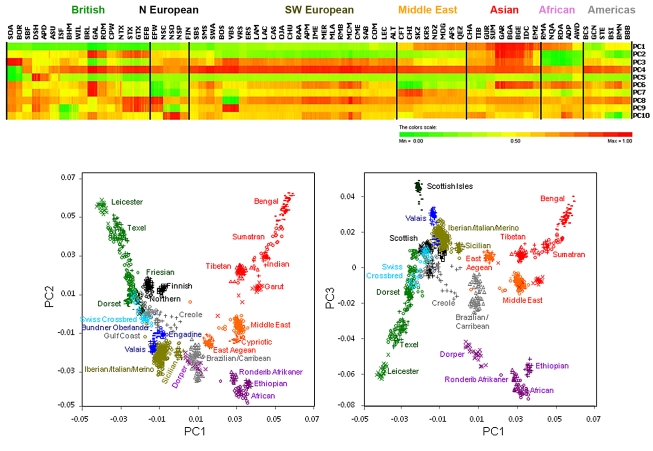
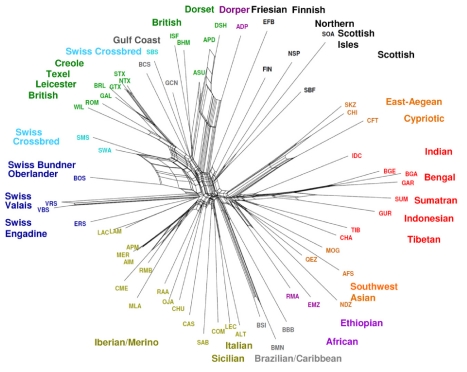
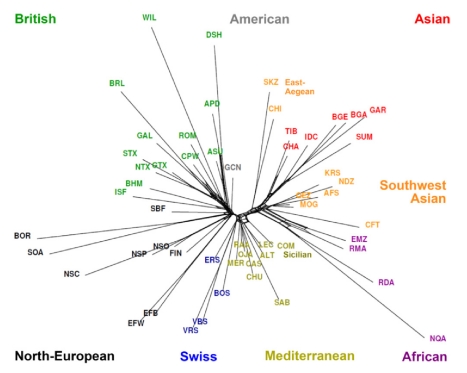
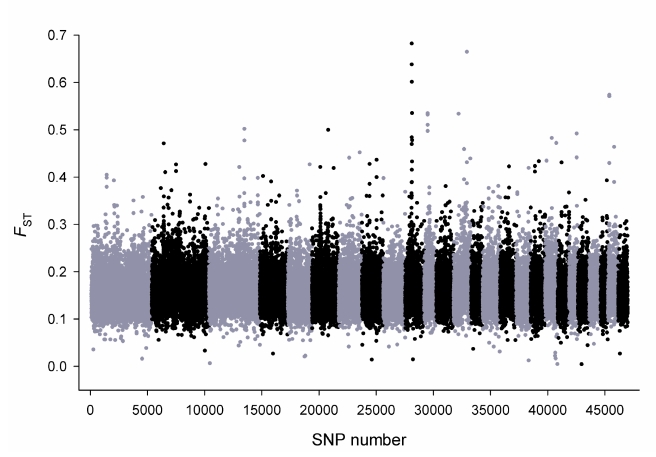
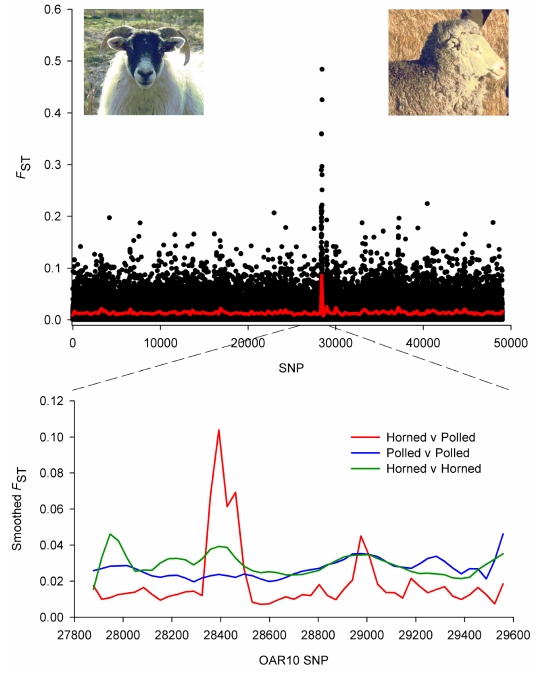
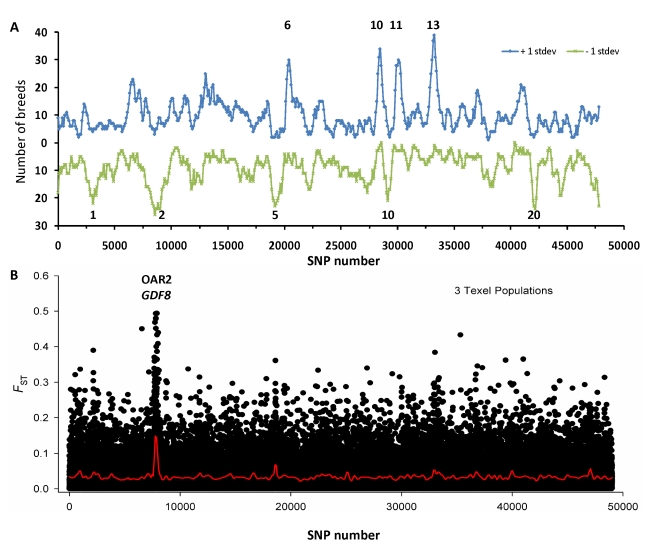
Through their domestication and subsequent selection, sheep have been adapted to thrive in a diverse range of environments. To characterise the genetic consequence of both domestication and selection, we genotyped 49,034 SNP in 2,819 animals from a diverse collection of 74 sheep breeds. We find the majority of sheep populations contain high SNP diversity and have retained an effective population size much higher than most cattle or dog breeds, suggesting domestication occurred from a broad genetic base. Extensive haplotype sharing and generally low divergence time between breeds reveal frequent genetic exchange has occurred during the development of modern breeds. A scan of the genome for selection signals revealed 31 regions containing genes for coat pigmentation, skeletal morphology, body size, growth, and reproduction. We demonstrate the strongest selection signal has occurred in response to breeding for the absence of horns. The high density map of genetic variability provides an in-depth view of the genetic history for this important livestock species.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Meadows J. R, Li K, Kantanen J, Tapio M, Sipos W, Pardeshi V, et al. Mitochondrial sequence reveals high levels of gene flow between breeds of domestic sheep from Asia and Europe. J Hered. 2005;96:494–501. - PubMed
-
- Tapio M, Marzanov N, Ozerov M, Cinkulov M, Gonzarenko G, et al. Sheep mitochondrial DNA variation in European, Caucasian, and Central Asian areas. Mol Biol Evol. 2006;23:1776–1783. - PubMed
-
- Tapio M, Tapio I, Grislis Z, Holm L. E, Jeppsson S, et al. Native breeds demonstrate high contributions to the molecular variation in northern European sheep. Mol Ecol. 2005;14:3951–3963. - PubMed
