

Structural and functional insights into endoglin ligand recognition and binding
- PMID: 22347366
- PMCID: PMC3275592
- DOI: 10.1371/journal.pone.0029948
Structural and functional insights into endoglin ligand recognition and binding
Abstract
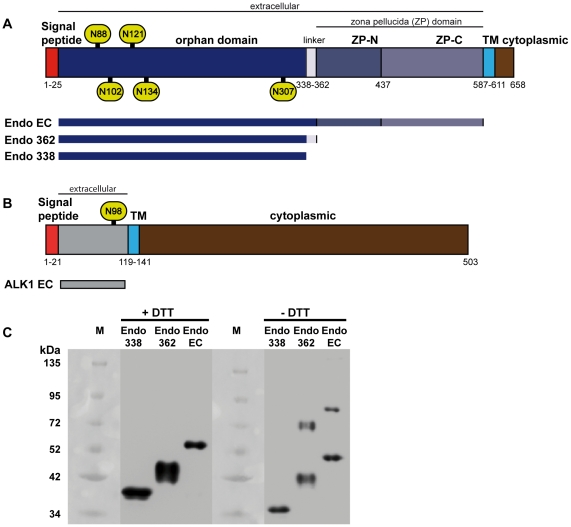
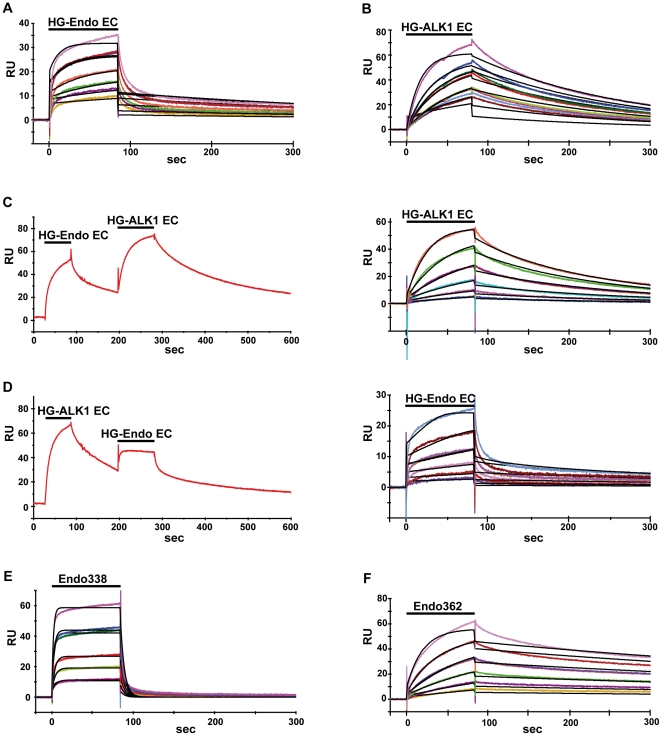
Endoglin, a type I membrane glycoprotein expressed as a disulfide-linked homodimer on human vascular endothelial cells, is a component of the transforming growth factor (TGF)-β receptor complex and is implicated in a dominant vascular dysplasia known as hereditary hemorrhagic telangiectasia as well as in preeclampsia. It interacts with the type I TGF-β signaling receptor activin receptor-like kinase (ALK)1 and modulates cellular responses to Bone Morphogenetic Protein (BMP)-9 and BMP-10. Structurally, besides carrying a zona pellucida (ZP) domain, endoglin contains at its N-terminal extracellular region a domain of unknown function and without homology to any other known protein, therefore called the orphan domain (OD). In this study, we have determined the recognition and binding ability of full length ALK1, endoglin and constructs encompassing the OD to BMP-9 using combined methods, consisting of surface plasmon resonance and cellular assays. ALK1 and endoglin ectodomains bind, independently of their glycosylation state and without cooperativity, to different sites of BMP-9. The OD comprising residues 22 to 337 was identified among the present constructs as the minimal active endoglin domain needed for partner recognition. These studies also pinpointed to Cys350 as being responsible for the dimerization of endoglin. In contrast to the complete endoglin ectodomain, the OD is a monomer and its small angle X-ray scattering characterization revealed a compact conformation in solution into which a de novo model was fitted.
Conflict of interest statement
Figures


References
-
- Bernabeu C, Lopez-Novoa JM, Quintanilla M. The emerging role of TGF-beta superfamily coreceptors in cancer. Biochim Biophys Acta. 2009;1792:954–973. - PubMed
-
- Lebrin F, Mummery CL. Endoglin-mediated vascular remodeling: mechanisms underlying hereditary hemorrhagic telangiectasia. Trends Cardiovasc Med. 2008;18:25–32. - PubMed
-
- Lopez-Novoa JM, Bernabeu C. The physiological role of endoglin in the cardiovascular system. Am J Physiol Heart Circ Physiol. 2010;299:H959–974. - PubMed
-
- McAllister KA, Grogg KM, Johnson DW, Gallione CJ, Baldwin MA, et al. Endoglin, a TGF-beta binding protein of endothelial cells, is the gene for hereditary haemorrhagic telangiectasia type 1. Nat Genet. 1994;8:345–351. - PubMed
-
- Bernabeu C, Blanco FJ, Langa C, Garrido-Martin EM, Botella LM. Involvement of the TGF-beta superfamily signalling pathway in hereditary haemorrhagic telangiectasia. J Appl Biomed. 2010;8:169–171.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
