

KSHV Rta Promoter Specification and Viral Reactivation
- PMID: 22347875
- PMCID: PMC3278982
- DOI: 10.3389/fmicb.2012.00030
KSHV Rta Promoter Specification and Viral Reactivation
Abstract
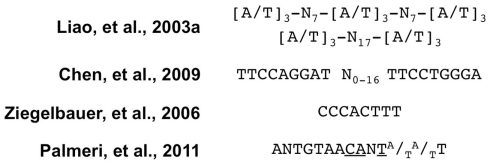
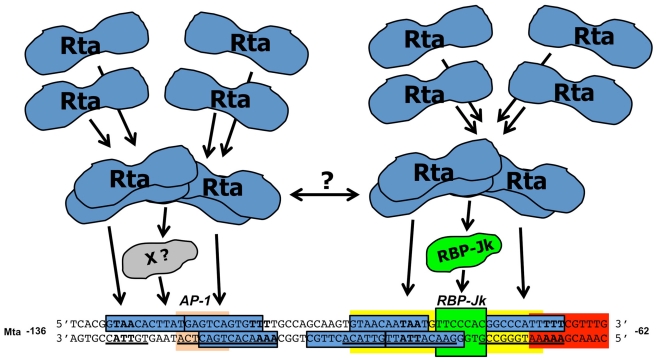
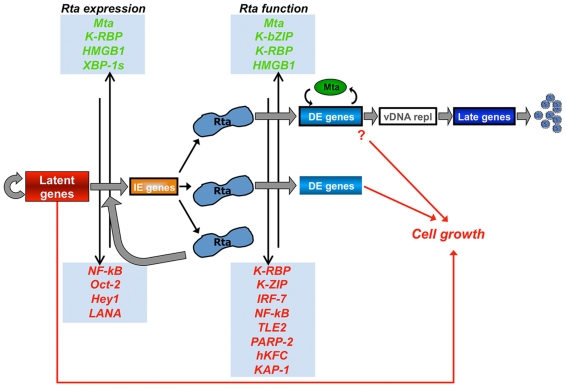
Viruses are obligate intracellular pathogens whose biological success depends upon replication and packaging of viral genomes, and transmission of progeny viruses to new hosts. The biological success of herpesviruses is enhanced by their ability to reproduce their genomes without producing progeny viruses or killing the host cells, a process called latency. Latency permits a herpesvirus to remain undetected in its animal host for decades while maintaining the potential to reactivate, or switch, to a productive life cycle when host conditions are conducive to generating viral progeny. Direct interactions between many host and viral molecules are implicated in controlling herpesviral reactivation, suggesting complex biological networks that control the decision. One viral protein that is necessary and sufficient to switch latent Kaposi's sarcoma-associated herpesvirus (KSHV) into the lytic infection cycle is called K-Rta. K-Rta is a transcriptional activator that specifies promoters by binding DNA directly and interacting with cellular proteins. Among these cellular proteins, binding of K-Rta to RBP-Jk is essential for viral reactivation. In contrast to the canonical model for Notch signaling, RBP-Jk is not uniformly and constitutively bound to the latent KSHV genome, but rather is recruited to DNA by interactions with K-Rta. Stimulation of RBP-Jk DNA binding requires high affinity binding of Rta to repetitive and palindromic "CANT DNA repeats" in promoters, and formation of ternary complexes with RBP-Jk. However, while K-Rta expression is necessary for initiating KSHV reactivation, K-Rta's role as the switch is inefficient. Many factors modulate K-Rta's function, suggesting that KSHV reactivation can be significantly regulated post-Rta expression and challenging the notion that herpesviral reactivation is bistable. This review analyzes rapidly evolving research on KSHV K-Rta to consider the role of K-Rta promoter specification in regulating the progression of KSHV reactivation.
Keywords: DNA binding; DNA/protein interactions; KSHV; RBP-Jk; Rta; herpesvirus; reactivation.
Figures



Similar articles
-
The cellular Notch1 protein promotes KSHV reactivation in an Rta-dependent manner.J Virol. 2024 Aug 20;98(8):e0078824. doi: 10.1128/jvi.00788-24. Epub 2024 Jul 8. J Virol. 2024. PMID: 38975769 Free PMC article.
-
Kaposi's sarcoma-associated herpesvirus Rta tetramers make high-affinity interactions with repetitive DNA elements in the Mta promoter to stimulate DNA binding of RBP-Jk/CSL.J Virol. 2011 Nov;85(22):11901-15. doi: 10.1128/JVI.05479-11. Epub 2011 Aug 31. J Virol. 2011. PMID: 21880753 Free PMC article.
-
Convergence of Kaposi's sarcoma-associated herpesvirus reactivation with Epstein-Barr virus latency and cellular growth mediated by the notch signaling pathway in coinfected cells.J Virol. 2010 Oct;84(20):10488-500. doi: 10.1128/JVI.00894-10. Epub 2010 Aug 4. J Virol. 2010. PMID: 20686042 Free PMC article.
-
KSHV and the Role of Notch Receptor Dysregulation in Disease Progression.Pathogens. 2017 Aug 4;6(3):34. doi: 10.3390/pathogens6030034. Pathogens. 2017. PMID: 28777778 Free PMC article. Review.
-
KSHV reactivation and novel implications of protein isomerization on lytic switch control.Viruses. 2015 Jan 12;7(1):72-109. doi: 10.3390/v7010072. Viruses. 2015. PMID: 25588053 Free PMC article. Review.
Cited by
-
Kaposi's sarcoma-associated herpesvirus replication and transcription activator protein activates CD274/PD-L1 gene promoter.Cancer Sci. 2023 Apr;114(4):1718-1728. doi: 10.1111/cas.15673. Epub 2022 Dec 16. Cancer Sci. 2023. PMID: 36411531 Free PMC article.
-
Molecular Biology of KSHV in Relation to HIV/AIDS-Associated Oncogenesis.Cancer Treat Res. 2019;177:23-62. doi: 10.1007/978-3-030-03502-0_2. Cancer Treat Res. 2019. PMID: 30523620 Free PMC article.
-
The regulation of KSHV lytic reactivation by viral and cellular factors.Curr Opin Virol. 2022 Feb;52:39-47. doi: 10.1016/j.coviro.2021.11.004. Epub 2021 Dec 3. Curr Opin Virol. 2022. PMID: 34872030 Free PMC article. Review.
-
KSHV: pathways to tumorigenesis and persistent infection.Adv Virus Res. 2014;88:111-59. doi: 10.1016/B978-0-12-800098-4.00002-7. Adv Virus Res. 2014. PMID: 24373311 Free PMC article. Review.
-
Egr-1 regulates RTA transcription through a cooperative involvement of transcriptional regulators.Oncotarget. 2017 Sep 5;8(53):91425-91444. doi: 10.18632/oncotarget.20648. eCollection 2017 Oct 31. Oncotarget. 2017. PMID: 29207655 Free PMC article.
References
-
- An F. Q., Folarin H. M., Compitello N., Roth J., Gerson S. L., Mccrae K. R., Fakhari F. D., Dittmer D. P., Renne R. (2006). Long-term-infected telomerase-immortalized endothelial cells: a model for Kaposi’s sarcoma-associated herpesvirus latency in vitro and in vivo. J. Virol. 80, 4833–484610.1128/JVI.80.10.4833-4846.2006 - DOI - PMC - PubMed
-
- AuCoin D. P., Colletti K. S., Cei S. A., Papouskova I., Tarrant M., Pari G. S. (2004). Amplification of the Kaposi’s sarcoma-associated herpesvirus/human herpesvirus 8 lytic origin of DNA replication is dependent upon a cis-acting AT-rich region and an ORF50 response element and the trans-acting factors ORF50 (K-Rta) and K8 (K-bZIP). Virology 318, 542–55510.1016/j.virol.2003.10.016 - DOI - PubMed
-
- Bello L., Davison A., Glenn M., Whitehouse A., Rethmeier N., Schulz T., Clements J. (1999). The human herpesvirus-8 ORF57 gene and its properties. J. Gen. Virol. 80, 3207–3215 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources