

Post-Translational Modifications of Kaposi's Sarcoma-Associated Herpesvirus Regulatory Proteins - SUMO and KSHV
- PMID: 22347876
- PMCID: PMC3278983
- DOI: 10.3389/fmicb.2012.00031
Post-Translational Modifications of Kaposi's Sarcoma-Associated Herpesvirus Regulatory Proteins - SUMO and KSHV
Abstract
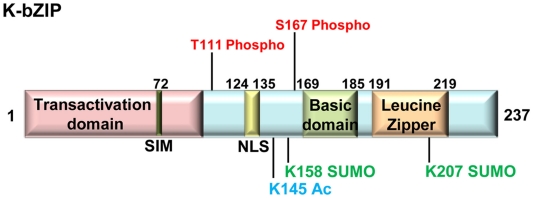
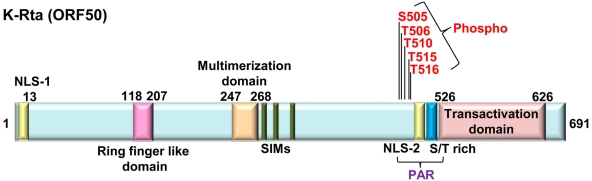
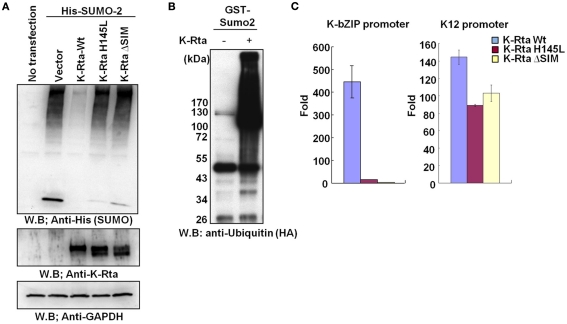
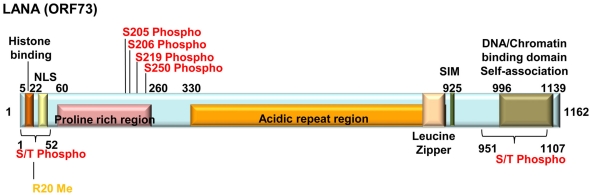
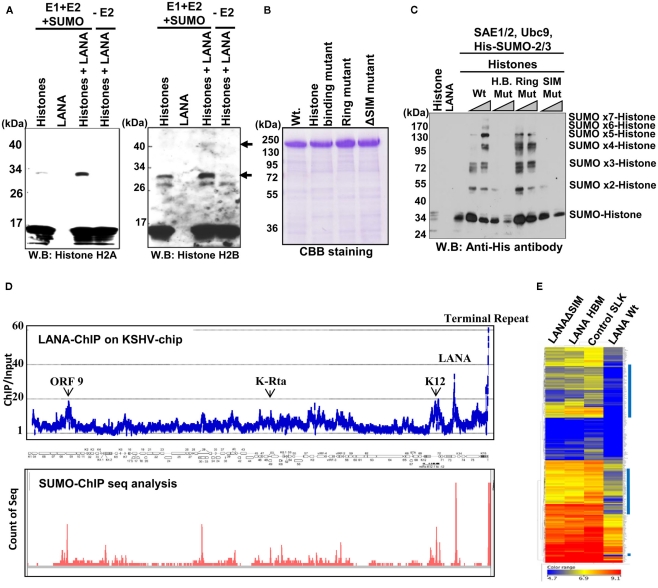
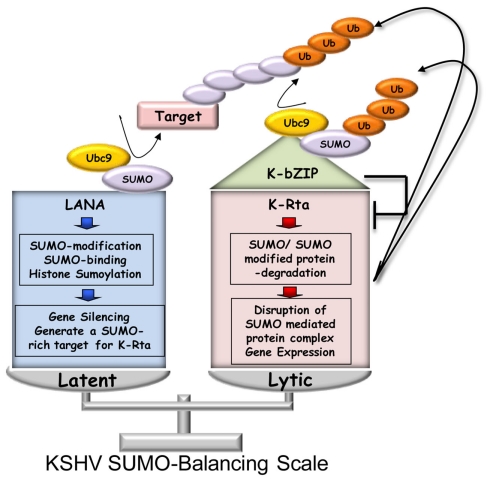
KSHV latency can be envisioned as an outcome that is balanced between factors that promote viral gene expression and lytic replication against those that facilitate gene silencing and establish or maintain latency. A large body of work has focused on the activities of the key viral regulatory proteins involved in KSHV latent or lytic states. Moreover, recent studies have also begun to document the importance of epigenetic landscape evolution of the KSHV viral genome during latency and reactivation. However, one area of KSHV molecular virology that remains largely unanswered is the precise role of post-translational modifications on the activities of viral factors that function during latency and reactivation. In this review, we will summarize the post-translational modifications associated with three viral factors whose activities contribute to the viral state. The viral proteins discussed are the two major KSHV encoded transcription factors, K-Rta (KSHV replication and transcriptional activator) and K-bZIP (KSHV basic leucine zipper) and the viral latency-associated nuclear antigen (LANA). A special emphasis will be placed on the role of the sumoylation pathway in the modulation of the KSHV lifecycle. Newly uncovered small ubiquitin-like modifier (SUMO)-associated properties of LANA and K-Rta will also be presented, namely LANA histone targeting SUMO E3 ligase activity and K-Rta SUMO-targeted ubiquitin ligase function.
Keywords: K-Rta; K-bZIP; KSHV; LANA; SUMO; post-translational modification; transcription.
Figures
References
-
- An F. Q., Compitello N., Horwitz E., Sramkoski M., Knudsen E. S., Renne R. (2005). The latency-associated nuclear antigen of Kaposi’s sarcoma-associated herpesvirus modulates cellular gene expression and protects lymphoid cells from p16 INK4A-induced cell cycle arrest. J. Biol. Chem. 280, 3862–3874 10.1074/jbc.M507550200 - DOI - PubMed