

Prevalence and genetic characterization of pertactin-deficient Bordetella pertussis in Japan
- PMID: 22348138
- PMCID: PMC3279416
- DOI: 10.1371/journal.pone.0031985
Prevalence and genetic characterization of pertactin-deficient Bordetella pertussis in Japan
Abstract
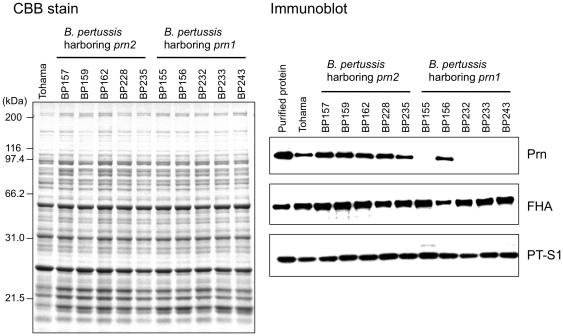
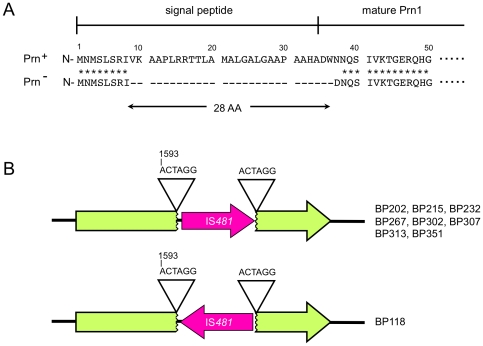
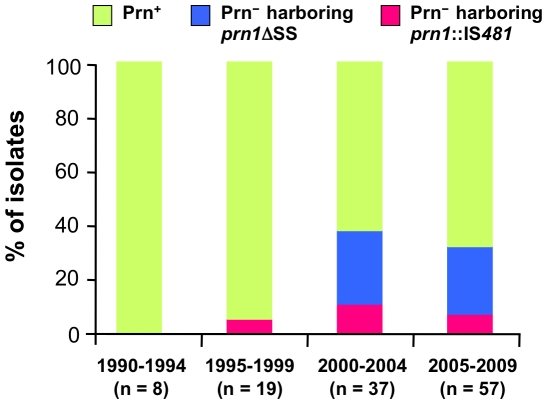
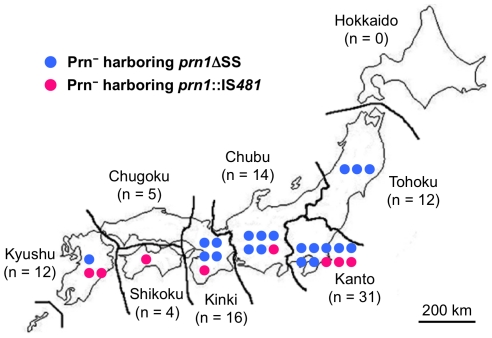
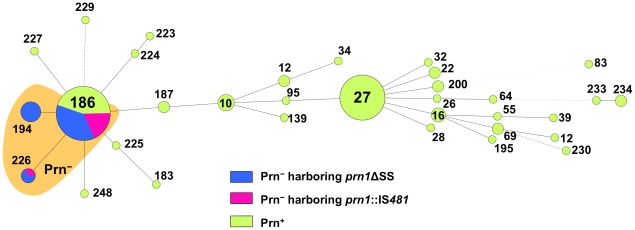
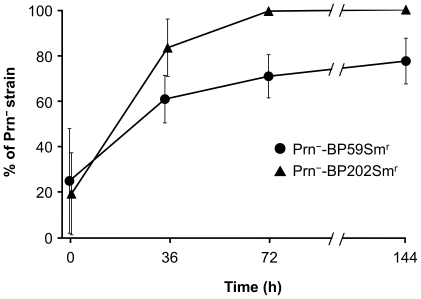
The adhesin pertactin (Prn) is one of the major virulence factors of Bordetella pertussis, the etiological agent of whooping cough. However, a significant prevalence of Prn-deficient (Prn(-)) B. pertussis was observed in Japan. The Prn(-) isolate was first discovered in 1997, and 33 (27%) Prn(-) isolates were identified among 121 B. pertussis isolates collected from 1990 to 2009. Sequence analysis revealed that all the Prn(-) isolates harbor exclusively the vaccine-type prn1 allele and that loss of Prn expression is caused by 2 different mutations: an 84-bp deletion of the prn signal sequence (prn1ΔSS, n = 24) and an IS481 insertion in prn1 (prn1::IS481, n = 9). The frequency of Prn(-) isolates, notably those harboring prn1ΔSS, significantly increased since the early 2000s, and Prn(-) isolates were subsequently found nationwide. Multilocus variable-number tandem repeat analysis (MLVA) revealed that 24 (73%) of 33 Prn(-) isolates belong to MLVA-186, and 6 and 3 Prn(-) isolates belong to MLVA-194 and MLVA-226, respectively. The 3 MLVA types are phylogenetically closely related, suggesting that the 2 Prn(-) clinical strains (harboring prn1ΔSS and prn1::IS481) have clonally expanded in Japan. Growth competition assays in vitro also demonstrated that Prn(-) isolates have a higher growth potential than the Prn(+) back-mutants from which they were derived. Our observations suggested that human host factors (genetic factors and immune status) that select for Prn(-) strains have arisen and that Prn expression is not essential for fitness under these conditions.
Conflict of interest statement
Figures
References
-
- von König CH, Halperin S, Riffelmann M, Guiso N. Pertussis of adults and infants. Lancet Infect Dis. 2002;2:744–750. - PubMed
-
- Hewlett EL, Edwards KM. Clinical practice. Pertussis–not just for kids. N Engl J Med. 2005;352:1215–1222. - PubMed
-
- Okada K, Komiya T, Yamamoto A, Takahashi M, Kamachi K, et al. Safe and effective booster immunization using DTaP in teenagers. Vaccine. 2010;28:7626–7633. - PubMed
-
- Cherry JD. Comparative efficacy of acellular pertussis vaccines: an analysis of recent trials. Pediatr Infect Dis J. 1997;16:S90–96. - PubMed
-
- Olin P, Rasmussen F, Gustafsson L, Hallander HO, Heijbel H. Randomised controlled trial of two-component, three-component, and five-component acellular pertussis vaccines compared with whole-cell pertussis vaccine. Lancet. 1997;350:1569–1577. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
