

Autosomal monoallelic expression in the mouse
- PMID: 22348269
- PMCID: PMC3334567
- DOI: 10.1186/gb-2012-13-2-r10
Autosomal monoallelic expression in the mouse
Abstract
Background: Random monoallelic expression defines an unusual class of genes displaying random choice for expression between the maternal and paternal alleles. Once established, the allele-specific expression pattern is stably maintained and mitotically inherited. Examples of random monoallelic genes include those found on the X-chromosome and a subset of autosomal genes, which have been most extensively studied in humans. Here, we report a genome-wide analysis of random monoallelic expression in the mouse. We used high density mouse genome polymorphism mapping arrays to assess allele-specific expression in clonal cell lines derived from heterozygous mouse strains.
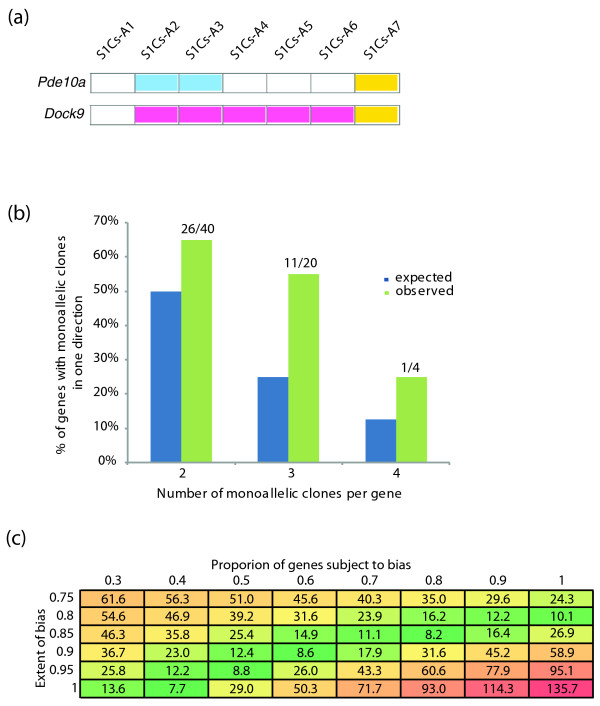
Results: Over 1,300 autosomal genes were assessed for allele-specific expression, and greater than 10% of them showed random monoallelic expression. When comparing mouse and human, the number of autosomal orthologs demonstrating random monoallelic expression in both organisms was greater than would be expected by chance. Random monoallelic expression on the mouse autosomes is broadly similar to that in human cells: it is widespread throughout the genome, lacks chromosome-wide coordination, and varies between cell types. However, for some mouse genes, there appears to be skewing, in some ways resembling skewed X-inactivation, wherein one allele is more frequently active.
Conclusions: These data suggest that autosomal random monoallelic expression was present at least as far back as the last common ancestor of rodents and primates. Random monoallelic expression can lead to phenotypic variation beyond the phenotypic variation dictated by genotypic variation. Thus, it is important to take into account random monoallelic expression when examining genotype-phenotype correlation.
Figures




Similar articles
-
Random monoallelic expression of genes on autosomes: Parallels with X-chromosome inactivation.Semin Cell Dev Biol. 2016 Aug;56:100-110. doi: 10.1016/j.semcdb.2016.04.007. Epub 2016 Apr 19. Semin Cell Dev Biol. 2016. PMID: 27101886 Review.
-
Widespread monoallelic expression on human autosomes.Science. 2007 Nov 16;318(5853):1136-40. doi: 10.1126/science.1148910. Science. 2007. PMID: 18006746
-
Allelic Imbalance Is a Prevalent and Tissue-Specific Feature of the Mouse Transcriptome.Genetics. 2015 Jun;200(2):537-49. doi: 10.1534/genetics.115.176263. Epub 2015 Apr 9. Genetics. 2015. PMID: 25858912 Free PMC article.
-
Single-cell RNA-seq reveals dynamic, random monoallelic gene expression in mammalian cells.Science. 2014 Jan 10;343(6167):193-6. doi: 10.1126/science.1245316. Science. 2014. PMID: 24408435
-
Random monoallelic expression: regulating gene expression one allele at a time.Trends Genet. 2014 Jun;30(6):237-44. doi: 10.1016/j.tig.2014.03.003. Epub 2014 Apr 26. Trends Genet. 2014. PMID: 24780084 Free PMC article. Review.
Cited by
-
Epigenetic regulation of monoallelic rearrangement (allelic exclusion) of antigen receptor genes.Front Immunol. 2014 Dec 5;5:625. doi: 10.3389/fimmu.2014.00625. eCollection 2014. Front Immunol. 2014. PMID: 25538709 Free PMC article. Review.
-
Random or stochastic monoallelic expressed genes are enriched for neurodevelopmental disorder candidate genes.PLoS One. 2013 Dec 27;8(12):e85093. doi: 10.1371/journal.pone.0085093. eCollection 2013. PLoS One. 2013. PMID: 24386451 Free PMC article.
-
The function of a heterozygous p53 mutation in a Li-Fraumeni syndrome patient.PLoS One. 2020 Jun 9;15(6):e0234262. doi: 10.1371/journal.pone.0234262. eCollection 2020. PLoS One. 2020. PMID: 32516327 Free PMC article.
-
Evolution of vertebrate sex chromosomes and dosage compensation.Nat Rev Genet. 2016 Jan;17(1):33-46. doi: 10.1038/nrg.2015.2. Epub 2015 Nov 30. Nat Rev Genet. 2016. PMID: 26616198 Review.
-
Genome-wide analysis of allele-specific expression of genes in the model diatom Phaeodactylum tricornutum.Sci Rep. 2021 Feb 3;11(1):2954. doi: 10.1038/s41598-021-82529-1. Sci Rep. 2021. PMID: 33536552 Free PMC article.
References
-
- Reik W, Walter J. Genomic imprinting: parental influence on the genome. Nat Rev Genet. 2001;2:21–32. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
