

Identification of novel pathways involved in the pathogenesis of human adamantinomatous craniopharyngioma
- PMID: 22349813
- PMCID: PMC3400760
- DOI: 10.1007/s00401-012-0957-9
Identification of novel pathways involved in the pathogenesis of human adamantinomatous craniopharyngioma
Abstract
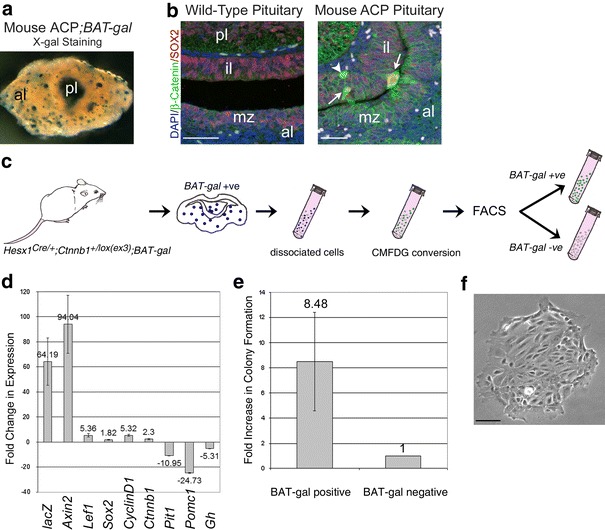
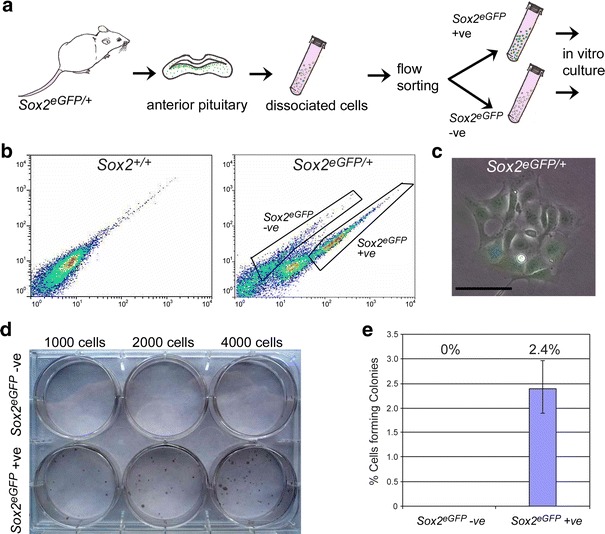
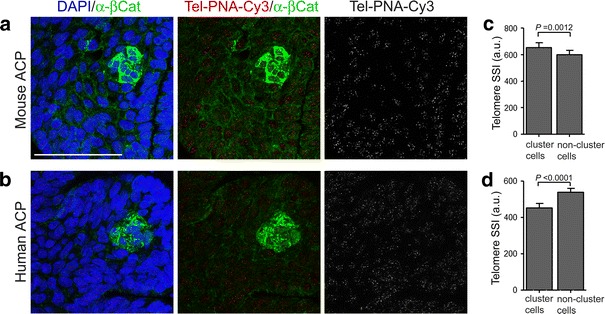
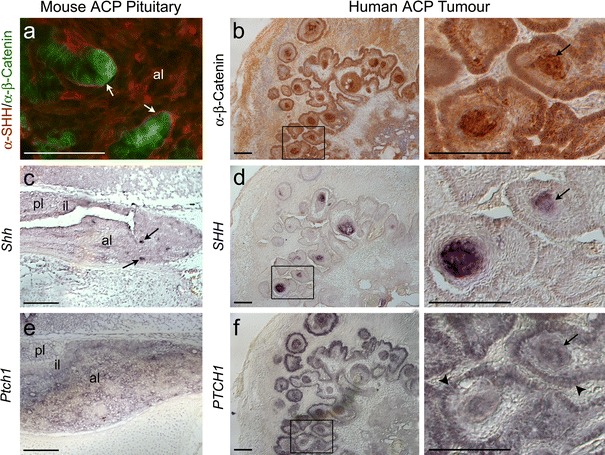
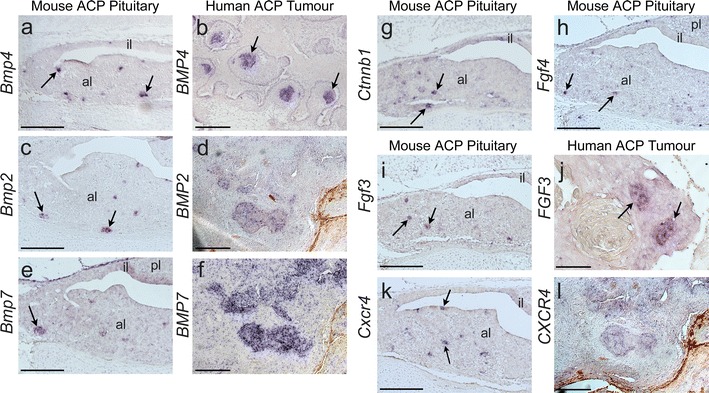
Activating mutations in the gene encoding β-catenin have been identified in the paediatric form of human craniopharyngioma (adamantinomatous craniopharyngioma, ACP), a histologically benign but aggressive pituitary tumour accounting for up to 10% of paediatric intracranial tumours. Recently, we generated an ACP mouse model and revealed that, as in human ACP, nucleocytoplasmic accumulation of β-catenin (β-cat(nc)) and over-activation of the Wnt/β-catenin pathway occurs only in a very small proportion of cells, which form clusters. Here, combining mouse genetics, fluorescence labelling and flow-sorting techniques, we have isolated these cells from tumorigenic mouse pituitaries and shown that the β-cat(nc) cells are enriched for colony-forming cells when cultured in stem cell-promoting media, and have longer telomeres, indicating shared properties with normal pituitary progenitors/stem cells (PSCs). Global gene profiling analysis has revealed that these β-cat(nc) cells express high levels of secreted mitogenic signals, such as members of the SHH, BMP and FGF family, in addition to several chemokines and their receptors, suggesting an important autocrine/paracrine role of these cells in the pathogenesis of ACP and a reciprocal communication with their environment. Finally, we highlight the clinical relevance of these findings by showing that these pathways are also up-regulated in the β-cat(nc) cell clusters identified in human ACP. As well as providing further support to the concept that pituitary stem cells may play an important role in the oncogenesis of human ACP, our data reveal novel disease biomarkers and potential pharmacological targets for the treatment of these devastating childhood tumours.
Figures
Similar articles
-
Molecular and cellular pathogenesis of adamantinomatous craniopharyngioma.Neuropathol Appl Neurobiol. 2015 Oct;41(6):721-32. doi: 10.1111/nan.12226. Epub 2015 Apr 23. Neuropathol Appl Neurobiol. 2015. PMID: 25611703 Free PMC article. Review.
-
60 YEARS OF NEUROENDOCRINOLOGY: Biology of human craniopharyngioma: lessons from mouse models.J Endocrinol. 2015 Aug;226(2):T161-72. doi: 10.1530/JOE-15-0145. Epub 2015 Apr 29. J Endocrinol. 2015. PMID: 25926515 Review.
-
Concise Review: Paracrine Role of Stem Cells in Pituitary Tumors: A Focus on Adamantinomatous Craniopharyngioma.Stem Cells. 2016 Feb;34(2):268-76. doi: 10.1002/stem.2267. Epub 2016 Jan 13. Stem Cells. 2016. PMID: 26763580 Free PMC article. Review.
-
Tumour compartment transcriptomics demonstrates the activation of inflammatory and odontogenic programmes in human adamantinomatous craniopharyngioma and identifies the MAPK/ERK pathway as a novel therapeutic target.Acta Neuropathol. 2018 May;135(5):757-777. doi: 10.1007/s00401-018-1830-2. Epub 2018 Mar 14. Acta Neuropathol. 2018. PMID: 29541918 Free PMC article.
-
Molecular pathology of adamantinomatous craniopharyngioma: review and opportunities for practice.Neurosurg Focus. 2016 Dec;41(6):E4. doi: 10.3171/2016.8.FOCUS16307. Neurosurg Focus. 2016. PMID: 27903120 Review.
Cited by
-
Inhibition of integrated stress response protects against lipid-induced senescence in hypothalamic neural stem cells in adamantinomatous craniopharyngioma.Neuro Oncol. 2023 Apr 6;25(4):720-732. doi: 10.1093/neuonc/noac261. Neuro Oncol. 2023. PMID: 36454228 Free PMC article.
-
SOX 2 Expression in Human Pituitary Adenomas-Correlations With Pituitary Function.In Vivo. 2019 Jan-Feb;33(1):79-83. doi: 10.21873/invivo.11442. In Vivo. 2019. PMID: 30587606 Free PMC article.
-
Homeostatic and tumourigenic activity of SOX2+ pituitary stem cells is controlled by the LATS/YAP/TAZ cascade.Elife. 2019 Mar 26;8:e43996. doi: 10.7554/eLife.43996. Elife. 2019. PMID: 30912742 Free PMC article.
-
Dynamic Expression of Imprinted Genes in the Developing and Postnatal Pituitary Gland.Genes (Basel). 2021 Mar 30;12(4):509. doi: 10.3390/genes12040509. Genes (Basel). 2021. PMID: 33808370 Free PMC article. Review.
-
FAP as a prognostic biomarker and radiomics-based predictor of angiogenesis-associated recurrence in Adamantinomatous craniopharyngioma.Pituitary. 2025 Jul 1;28(4):80. doi: 10.1007/s11102-025-01552-x. Pituitary. 2025. PMID: 40591015
References
-
- Andoniadou CL, Signore M, Sajedi E, Gaston-Massuet C, Kelberman D, Burns AJ, Itasaki N, Dattani M, Martinez-Barbera JP. Lack of the murine homeobox gene Hesx1 leads to a posterior transformation of the anterior forebrain. Development. 2007;134(8):1499–1508. doi: 10.1242/dev.02829. - DOI - PMC - PubMed
-
- Andoniadou CL, Signore M, Young RM, Gaston-Massuet C, Wilson SW, Fuchs E, Martinez-Barbera JP. HESX1- and TCF3-mediated repression of Wnt/beta-catenin targets is required for normal development of the anterior forebrain. Development. 2011;138(22):4931–4942. doi: 10.1242/dev.066597. - DOI - PMC - PubMed
-
- Barbieri F, Bajetto A, Stumm R, Pattarozzi A, Porcile C, Zona G, Dorcaratto A, Ravetti JL, Minuto F, Spaziante R, Schettini G, Ferone D, Florio T. Overexpression of stromal cell-derived factor 1 and its receptor CXCR4 induces autocrine/paracrine cell proliferation in human pituitary adenomas. Clin Cancer Res. 2008;14(16):5022–5032. doi: 10.1158/1078-0432.CCR-07-4717. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
