

Ligation of the fibrin-binding domain by β-strand addition is sufficient for expansion of soluble fibronectin
- PMID: 22351755
- PMCID: PMC3339936
- DOI: 10.1074/jbc.M111.294041
Ligation of the fibrin-binding domain by β-strand addition is sufficient for expansion of soluble fibronectin
Abstract
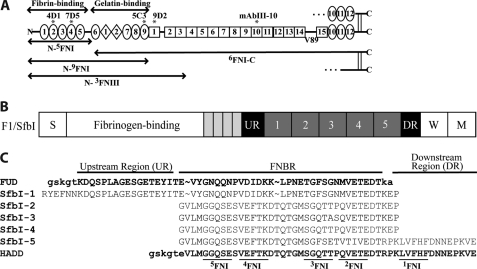
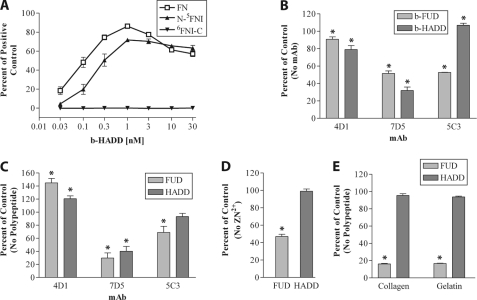
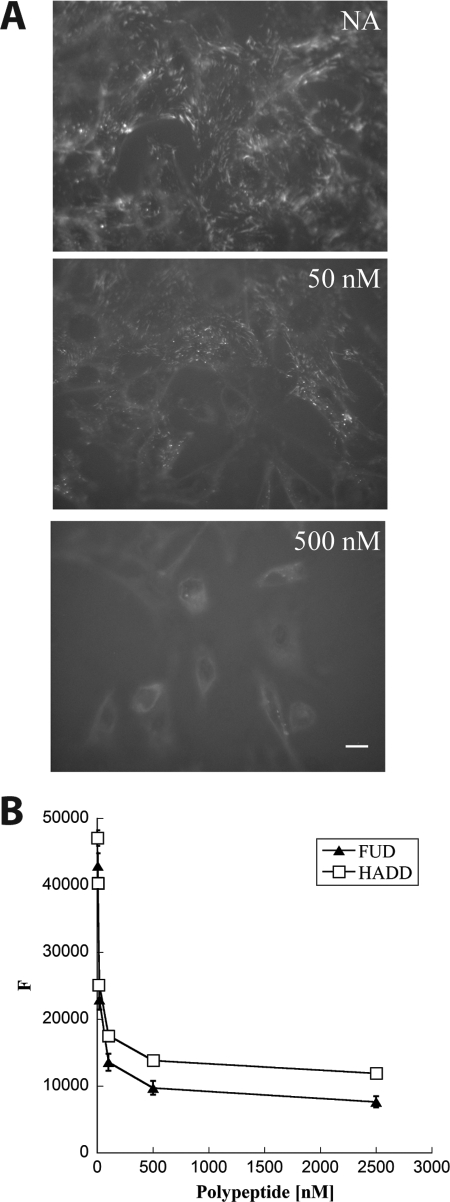
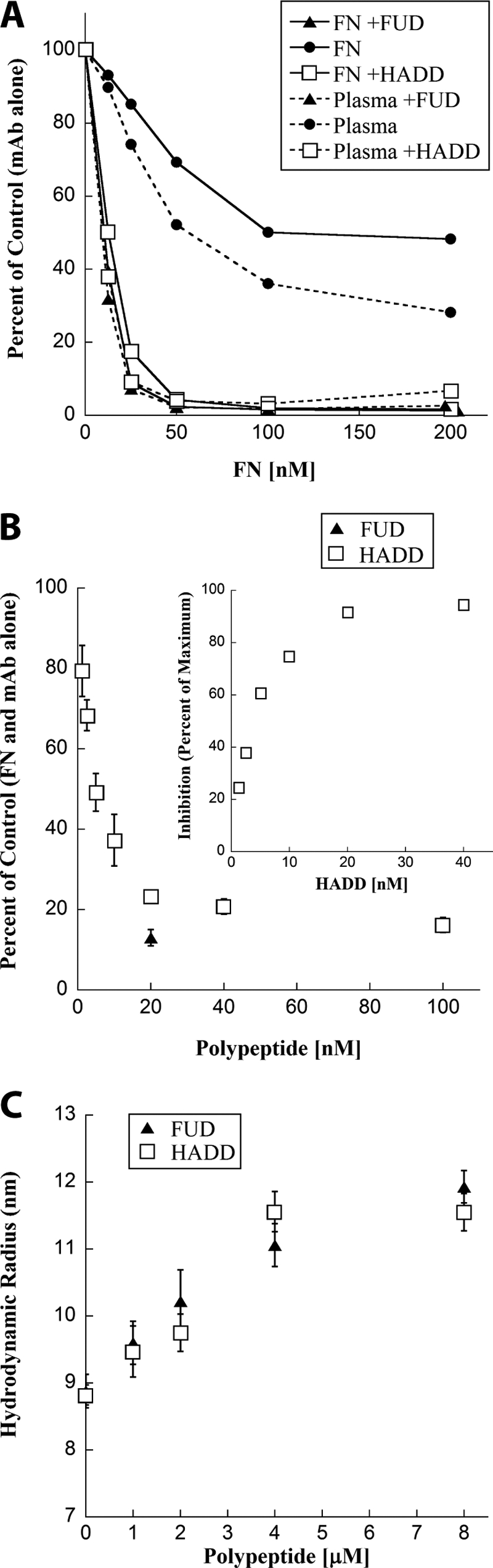
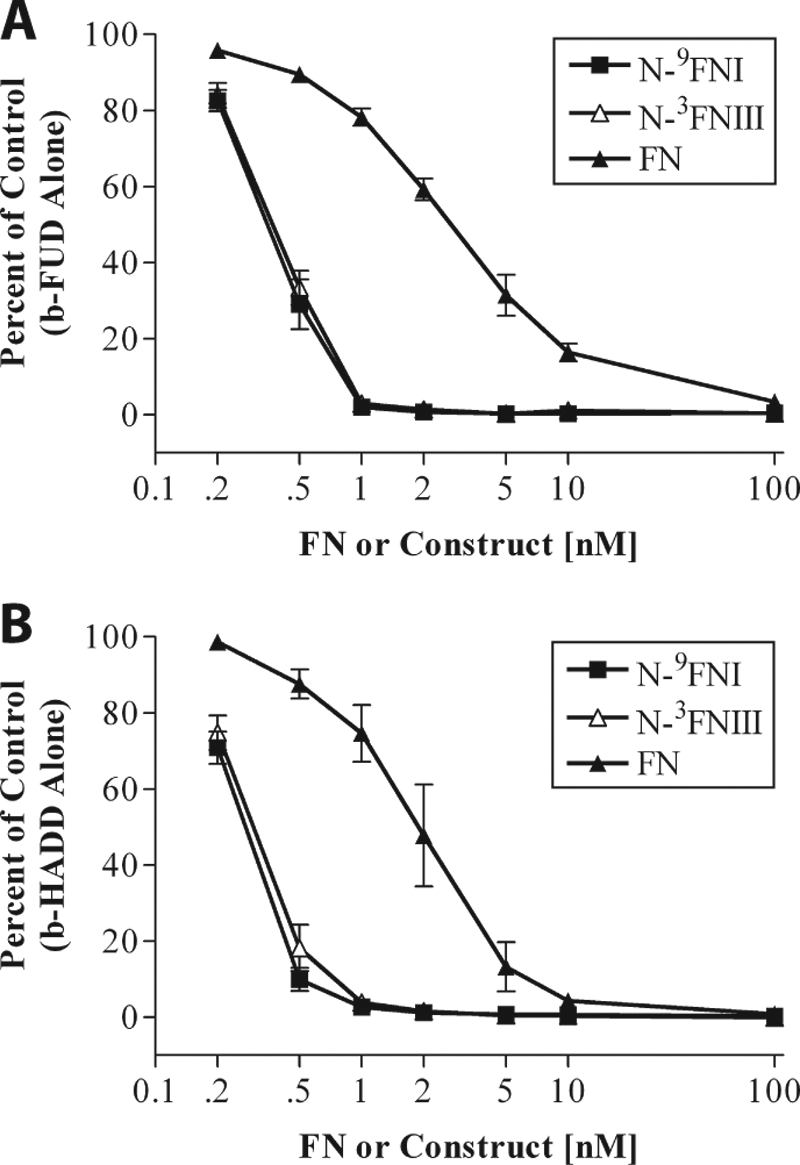
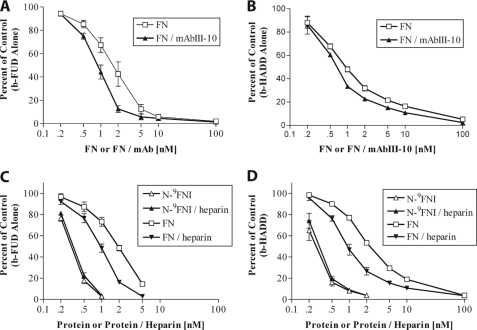
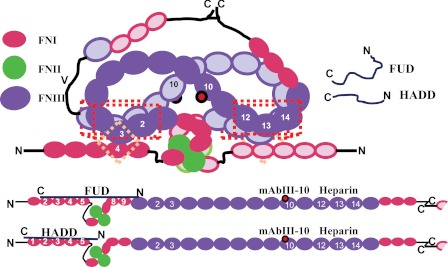
How fibronectin (FN) converts from a compact plasma protein to a fibrillar component of extracellular matrix is not understood. "Functional upstream domain" (FUD), a polypeptide based on F1 adhesin of Streptococcus pyogenes, binds by anti-parallel β-strand addition to discontinuous sets of N-terminal FN type I modules, (2-5)FNI of the fibrin-binding domain and (8-9)FNI of the gelatin-binding domain. Such binding blocks assembly of FN. To learn whether ligation of (2-5)FNI, (8-9)FNI, or the two sets in combination is important for inhibition, we tested "high affinity downstream domain" (HADD), which binds by β-strand addition to the continuous set of FNI modules, (1-5)FNI, comprising the fibrin-binding domain. HADD and FUD were similarly active in blocking fibronectin assembly. Binding of HADD or FUD to soluble plasma FN exposed the epitope to monoclonal antibody mAbIII-10 in the tenth FN type III module ((10)FNIII) and caused expansion of FN as assessed by dynamic light scattering. Soluble N-terminal constructs truncated after (9)FNI or (3)FNIII competed better than soluble FN for binding of FUD or HADD to adsorbed FN, indicating that interactions involving type III modules more C-terminal than (3)FNIII limit β-strand addition to (1-5)FNI within intact soluble FN. Preincubation of FN with mAbIII-10 or heparin modestly increased binding to HADD or FUD. Thus, ligation of FNIII modules involved in binding of integrins and glycosaminoglycans, (10)FNIII and (12-14)FNIII, increases accessibility of (1-5)FNI. Allosteric loss of constraining interactions among (1-5)FNI, (10)FNIII, and (12-14)FNIII likely enables assembly of FN into extracellular fibrils.
Figures
Similar articles
-
On-Off Kinetics of Engagement of FNI Modules of Soluble Fibronectin by β-Strand Addition.PLoS One. 2015 Apr 28;10(4):e0124941. doi: 10.1371/journal.pone.0124941. eCollection 2015. PLoS One. 2015. PMID: 25919138 Free PMC article.
-
Extended binding site on fibronectin for the functional upstream domain of protein F1 of Streptococcus pyogenes.J Biol Chem. 2010 Dec 24;285(52):41087-99. doi: 10.1074/jbc.M110.153692. Epub 2010 Oct 13. J Biol Chem. 2010. PMID: 20947497 Free PMC article.
-
IGD motifs, which are required for migration stimulatory activity of fibronectin type I modules, do not mediate binding in matrix assembly.PLoS One. 2012;7(2):e30615. doi: 10.1371/journal.pone.0030615. Epub 2012 Feb 15. PLoS One. 2012. PMID: 22355321 Free PMC article.
-
Pleiotropic virulence factor - Streptococcus pyogenes fibronectin-binding proteins.Cell Microbiol. 2013 Apr;15(4):503-11. doi: 10.1111/cmi.12083. Epub 2012 Dec 20. Cell Microbiol. 2013. PMID: 23190012 Review.
-
Roles of integrins in fibronectin matrix assembly.Histol Histopathol. 1997 Jan;12(1):233-40. Histol Histopathol. 1997. PMID: 9046058 Review.
Cited by
-
Fetal fibronectin signaling induces matrix metalloproteases and cyclooxygenase-2 (COX-2) in amnion cells and preterm birth in mice.J Biol Chem. 2013 Jan 18;288(3):1953-66. doi: 10.1074/jbc.M112.424366. Epub 2012 Nov 26. J Biol Chem. 2013. PMID: 23184961 Free PMC article.
-
Multiple Cryptic Binding Sites are Necessary for Robust Fibronectin Assembly: An In Silico Study.Sci Rep. 2017 Dec 22;7(1):18061. doi: 10.1038/s41598-017-18328-4. Sci Rep. 2017. PMID: 29273802 Free PMC article.
-
[64Cu]Cu-PEG-FUD peptide for noninvasive and sensitive detection of murine pulmonary fibrosis.Sci Adv. 2024 Apr 12;10(15):eadj1444. doi: 10.1126/sciadv.adj1444. Epub 2024 Apr 10. Sci Adv. 2024. PMID: 38598637 Free PMC article.
-
On-Off Kinetics of Engagement of FNI Modules of Soluble Fibronectin by β-Strand Addition.PLoS One. 2015 Apr 28;10(4):e0124941. doi: 10.1371/journal.pone.0124941. eCollection 2015. PLoS One. 2015. PMID: 25919138 Free PMC article.
-
The fibronectin-targeting PEG-FUD imaging probe shows enhanced uptake during fibrogenesis in experimental lung fibrosis.Respir Res. 2025 Jan 22;26(1):34. doi: 10.1186/s12931-025-03107-x. Respir Res. 2025. PMID: 39844185 Free PMC article.
References
-
- Pankov R., Yamada K. M. (2002) Fibronectin at a glance. J. Cell Sci. 115, 3861–3863 - PubMed
-
- Tucker R. P., Chiquet-Ehrismann R. (2009) Evidence for the evolution of tenascin and fibronectin early in the chordate lineage. Int. J. Biochem. Cell Biol. 41, 424–434 - PubMed
-
- Odermatt E., Engel J. (1989) Physical Properties of Fibronectin in Fibronectin (Mosher D. F., ed) pp. 29–45, Academic Press, Inc., San Diego, CA
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
