

CutA divalent cation tolerance homolog (Escherichia coli) (CUTA) regulates β-cleavage of β-amyloid precursor protein (APP) through interacting with β-site APP cleaving protein 1 (BACE1)
- PMID: 22351782
- PMCID: PMC3322838
- DOI: 10.1074/jbc.M111.330209
CutA divalent cation tolerance homolog (Escherichia coli) (CUTA) regulates β-cleavage of β-amyloid precursor protein (APP) through interacting with β-site APP cleaving protein 1 (BACE1)
Abstract
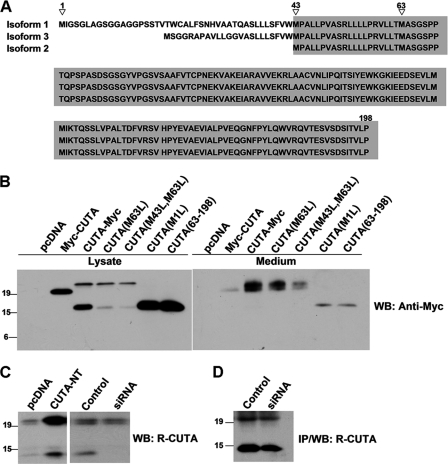
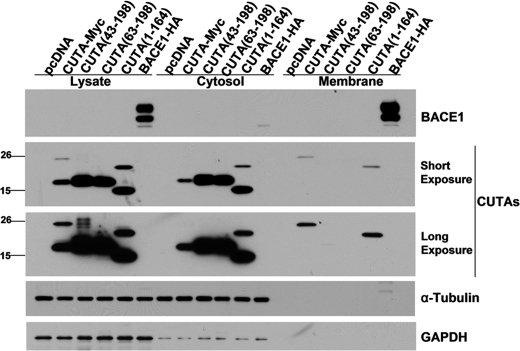
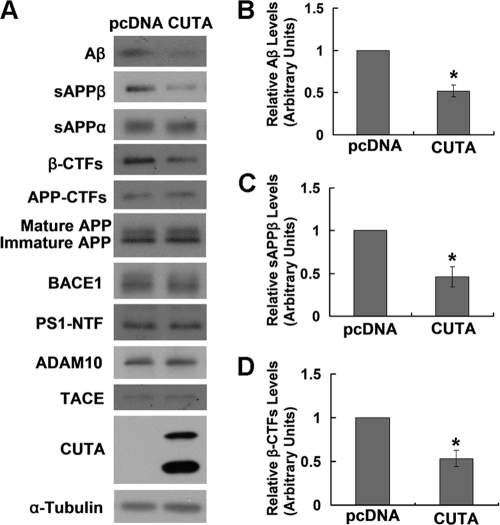
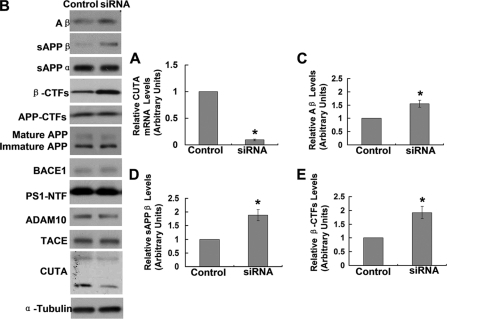
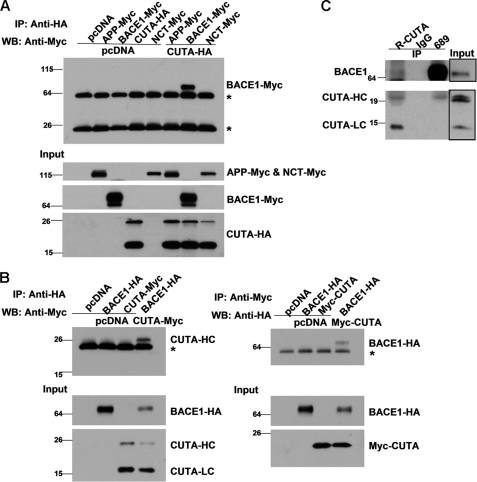
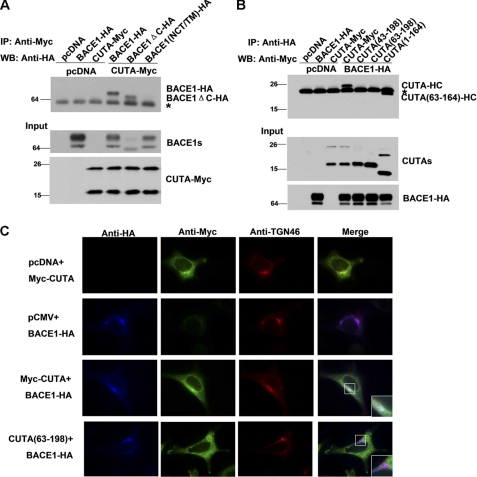
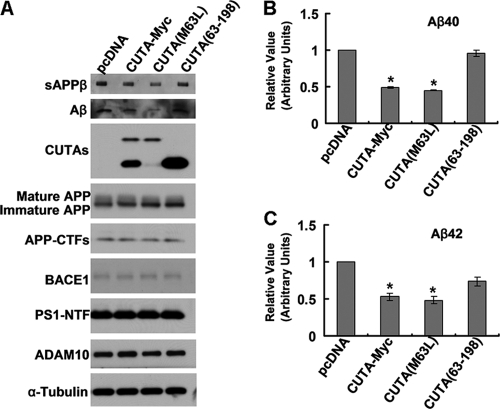
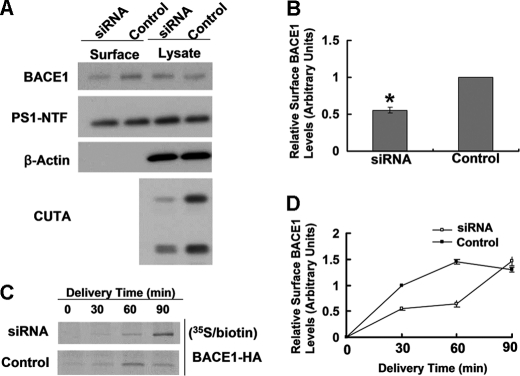
Accumulation of the neurotoxic β-amyloid (Aβ) peptide in the brain is central to the pathogenesis of Alzheimer disease. Aβ is derived from the β-amyloid precursor protein (APP) through sequential cleavages by β- and γ-secretases, and the production of Aβ is greatly affected by the subcellular localization of these factors. CUTA, the mammalian CutA divalent cation tolerance homolog (E. coli), has been proposed to mediate acetylcholinesterase activity and copper homeostasis, which are important in Alzheimer disease pathology. However, the exact function of CUTA remains largely unclear. Here we show that human CUTA has several variants that differ in their N-terminal length and are separated as heavy (H) and light (L) components. The H component has the longest N terminus and is membrane-associated, whereas the L component is N-terminally truncated at various sites and localized in the cytosol. Importantly, we demonstrate that the H component of CUTA interacts through its N terminus with the transmembrane domain of β-site APP cleaving enzyme 1 (BACE1), the putative β-secretase, mainly in the Golgi/trans-Golgi network. Overexpression and RNA interference knockdown of CUTA can reduce and increase BACE1-mediated APP processing/Aβ secretion, respectively. RNA interference of CUTA decelerates intracellular trafficking of BACE1 from the Golgi/trans-Golgi network to the cell surface and reduces the steady-state level of cell surface BACE1. Our results identify the H component of CUTA as a novel BACE1-interacting protein that mediates the intracellular trafficking of BACE1 and the processing of APP to Aβ.
Figures
References
-
- Glenner G. G., Wong C. W. (1984) Alzheimer disease. Initial report of the purification and characterization of a novel cerebrovascular amyloid protein. Biochem. Biophys. Res. Commun. 120, 885–890 - PubMed
-
- Buée L., Bussière T., Buée-Scherrer V., Delacourte A., Hof P. R. (2000) Tau protein isoforms, phosphorylation, and role in neurodegenerative disorders. Brain Res. Brain Res. Rev. 33, 95–130 - PubMed
-
- Sinha S., Anderson J. P., Barbour R., Basi G. S., Caccavello R., Davis D., Doan M., Dovey H. F., Frigon N., Hong J., Jacobson-Croak K., Jewett N., Keim P., Knops J., Lieberburg I., Power M., Tan H., Tatsuno G., Tung J., Schenk D., Seubert P., Suomensaari S. M., Wang S., Walker D., Zhao J., McConlogue L., John V. (1999) Purification and cloning of amyloid precursor protein β-secretase from human brain. Nature 402, 537–540 - PubMed
-
- Vassar R., Bennett B. D., Babu-Khan S., Kahn S., Mendiaz E. A., Denis P., Teplow D. B., Ross S., Amarante P., Loeloff R., Luo Y., Fisher S., Fuller J., Edenson S., Lile J., Jarosinski M. A., Biere A. L., Curran E., Burgess T., Louis J. C., Collins F., Treanor J., Rogers G., Citron M. (1999) β-Secretase cleavage of Alzheimer amyloid precursor protein by the transmembrane aspartic protease BACE. Science 286, 735–741 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
