

Prunasin hydrolases during fruit development in sweet and bitter almonds
- PMID: 22353576
- PMCID: PMC3320195
- DOI: 10.1104/pp.111.192021
Prunasin hydrolases during fruit development in sweet and bitter almonds
Abstract
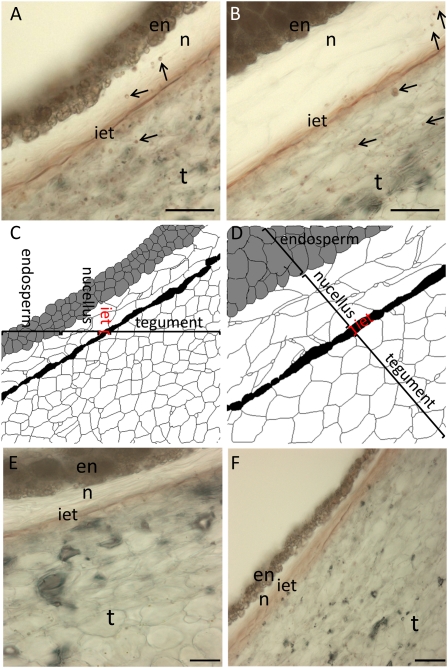
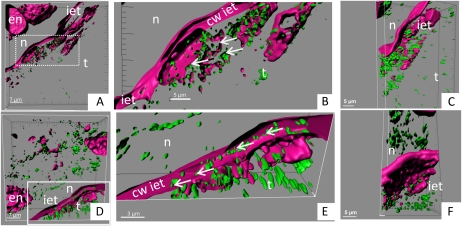
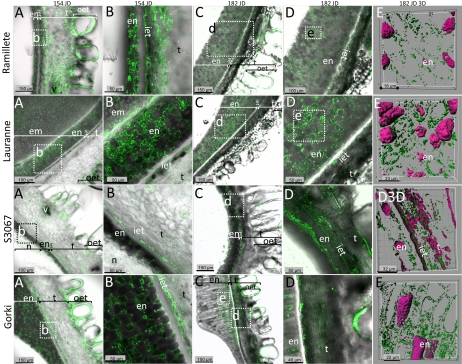
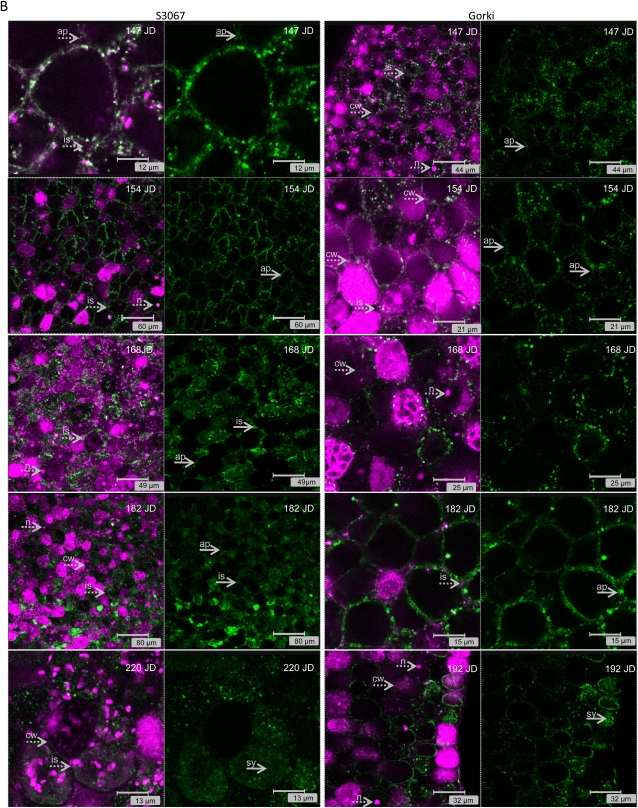
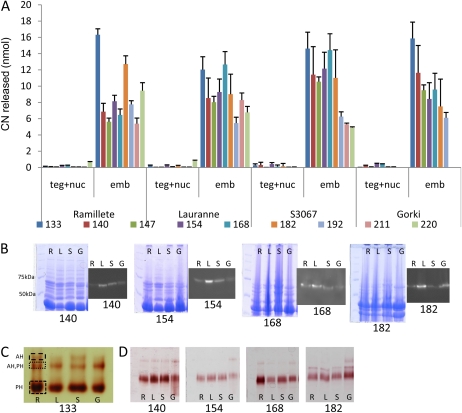
Amygdalin is a cyanogenic diglucoside and constitutes the bitter component in bitter almond (Prunus dulcis). Amygdalin concentration increases in the course of fruit formation. The monoglucoside prunasin is the precursor of amygdalin. Prunasin may be degraded to hydrogen cyanide, glucose, and benzaldehyde by the action of the β-glucosidase prunasin hydrolase (PH) and mandelonitirile lyase or be glucosylated to form amygdalin. The tissue and cellular localization of PHs was determined during fruit development in two sweet and two bitter almond cultivars using a specific antibody toward PHs. Confocal studies on sections of tegument, nucellus, endosperm, and embryo showed that the localization of the PH proteins is dependent on the stage of fruit development, shifting between apoplast and symplast in opposite patterns in sweet and bitter cultivars. Two different PH genes, Ph691 and Ph692, have been identified in a sweet and a bitter almond cultivar. Both cDNAs are 86% identical on the nucleotide level, and their encoded proteins are 79% identical to each other. In addition, Ph691 and Ph692 display 92% and 86% nucleotide identity to Ph1 from black cherry (Prunus serotina). Both proteins were predicted to contain an amino-terminal signal peptide, with the size of 26 amino acid residues for PH691 and 22 residues for PH692. The PH activity and the localization of the respective proteins in vivo differ between cultivars. This implies that there might be different concentrations of prunasin available in the seed for amygdalin synthesis and that these differences may determine whether the mature almond develops into bitter or sweet.
Figures


References
-
- Arrázola G. (2002) Análisis de glucósidos cianogénicos en variedades de almendro: implicaciones en la mejora genética. PhD thesis. Universidad de Alicante, Alicante, Spain
-
- Conn EE. (1980) Cyanogenic compounds. Annu Rev Plant Physiol 31: 433–451
-
- Dicenta F, Martínez-Gómez P, Grané N, Martín ML, León A, Cánovas JA, Berenguer V. (2002) Relationship between cyanogenic compounds in kernels, leaves, and roots of sweet and bitter kernelled almonds. J Agric Food Chem 50: 2149–2152 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
