

N-cadherin specifies first asymmetry in developing neurons
- PMID: 22354041
- PMCID: PMC3343329
- DOI: 10.1038/emboj.2012.41
N-cadherin specifies first asymmetry in developing neurons
Abstract
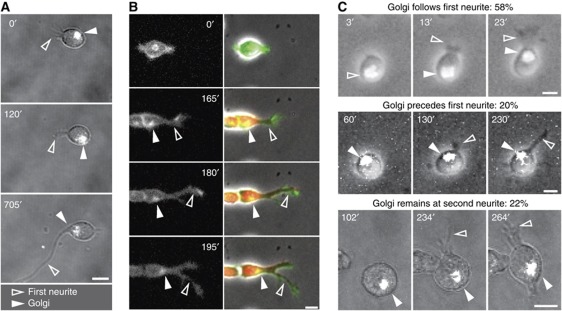
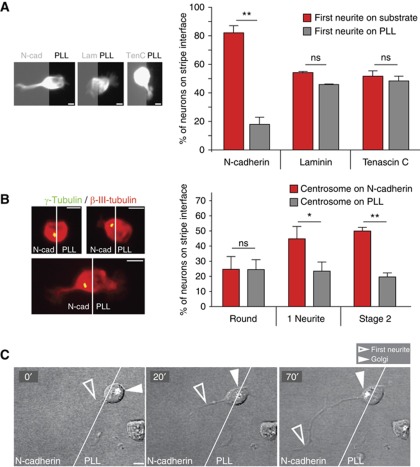
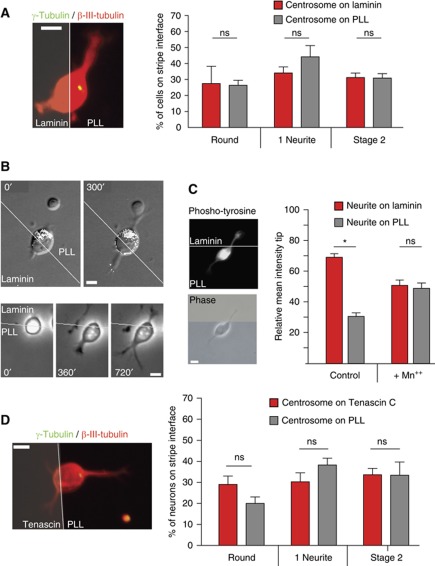
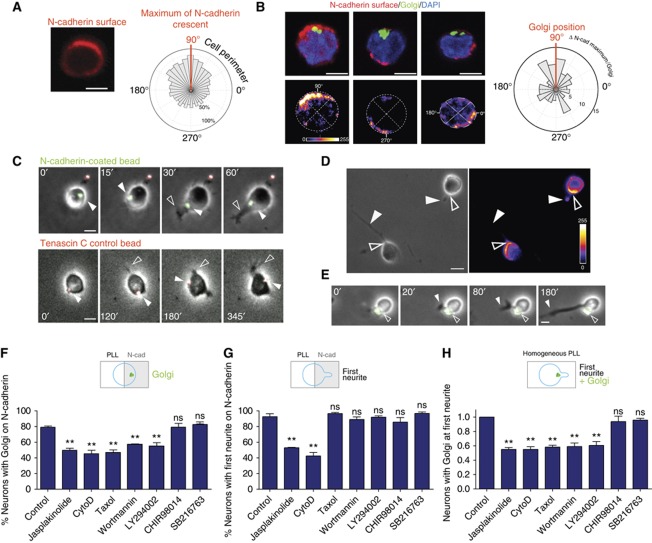
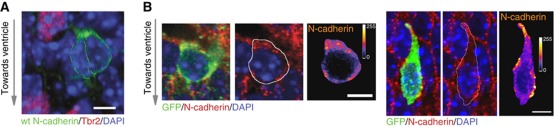
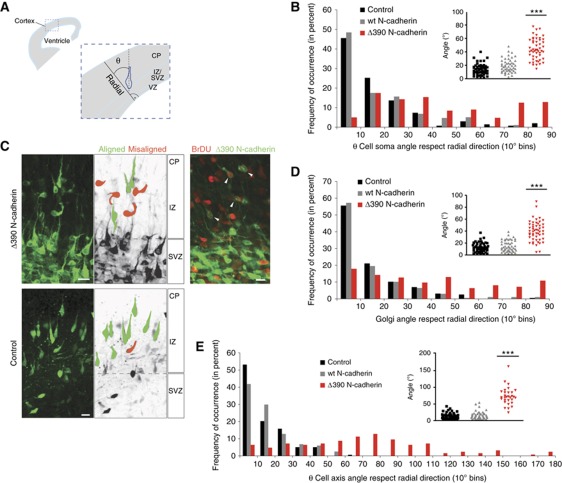
The precise polarization and orientation of developing neurons is essential for the correct wiring of the brain. In pyramidal excitatory neurons, polarization begins with the sprouting of opposite neurites, which later define directed migration and axo-dendritic domains. We here show that endogenous N-cadherin concentrates at one pole of the newborn neuron, from where the first neurite subsequently emerges. Ectopic N-cadherin is sufficient to favour the place of appearance of the first neurite. The Golgi and centrosome move towards this newly formed morphological pole in a second step, which is regulated by PI3K and the actin/microtubule cytoskeleton. Moreover, loss of function experiments in vivo showed that developing neurons with a non-functional N-cadherin misorient their cell axis. These results show that polarization of N-cadherin in the immediate post-mitotic stage is an early and crucial mechanism in neuronal polarity.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Banker G, Goslin K (1988) Developments in neuronal cell culture. Nature 336: 185–186 - PubMed
-
- Basto R, Lau J, Vinogradova T, Gardiol A, Woods CG, Khodjakov A, Raff JW (2006) Flies without centrioles. Cell 125: 1375–1386 - PubMed
-
- Chausovsky A, Bershadsky AD, Borisy GG (2000) Cadherin-mediated regulation of microtubule dynamics. Nat Cell Biol 2: 797–804 - PubMed
-
- Cowan CR, Hyman AA (2004) Centrosomes direct cell polarity independently of microtubule assembly in C. elegans embryos. Nature 431: 92–96 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
