

Popeye domain containing proteins are essential for stress-mediated modulation of cardiac pacemaking in mice
- PMID: 22354168
- PMCID: PMC3287222
- DOI: 10.1172/JCI59410
Popeye domain containing proteins are essential for stress-mediated modulation of cardiac pacemaking in mice
Abstract
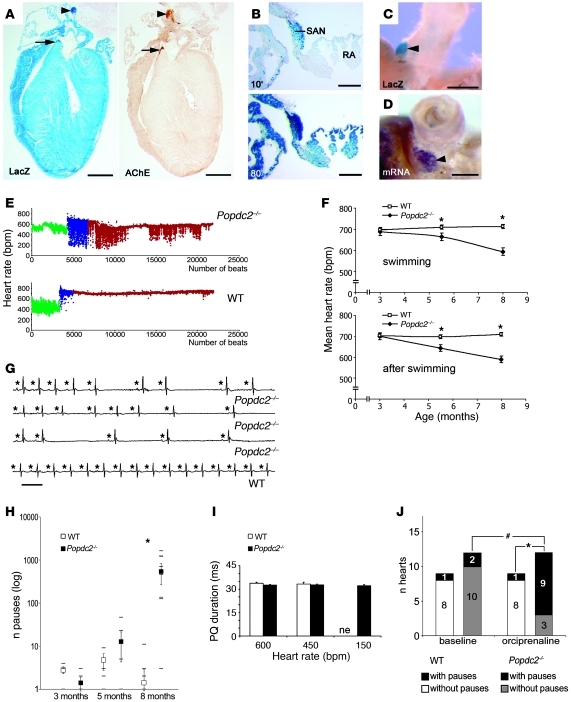
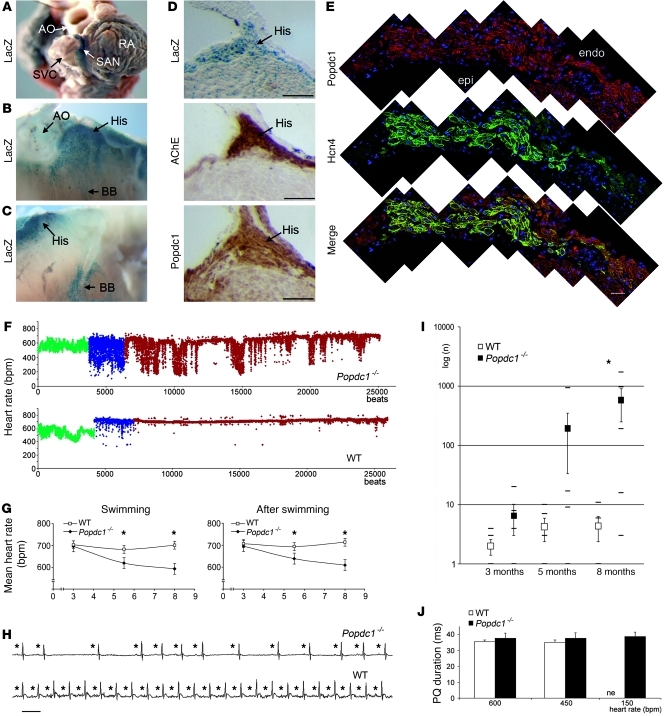
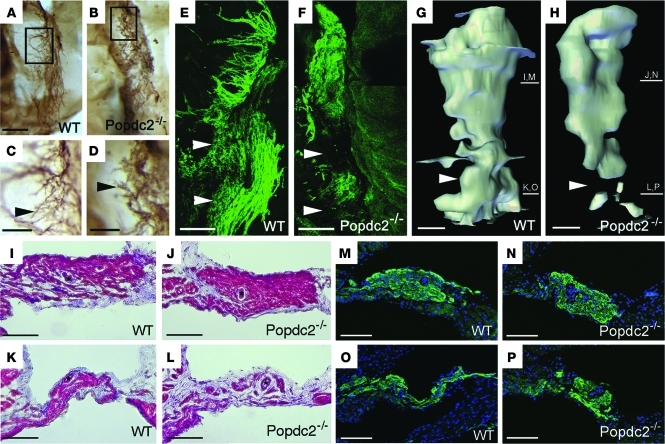
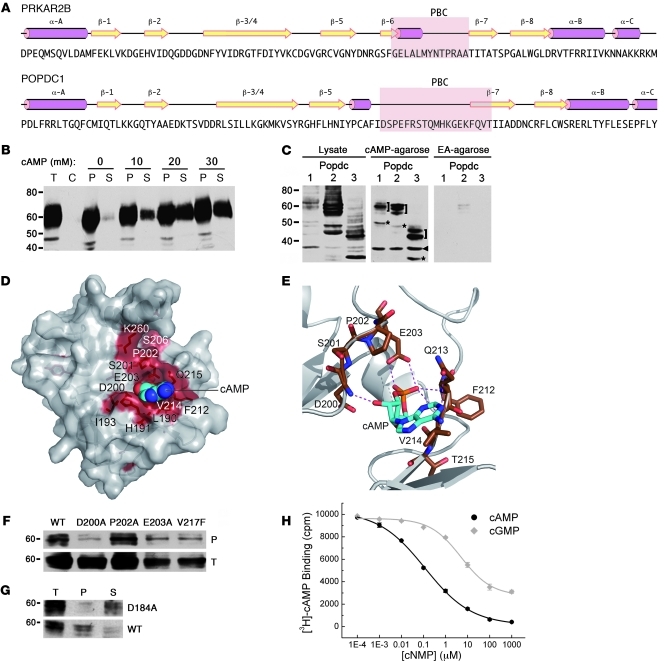
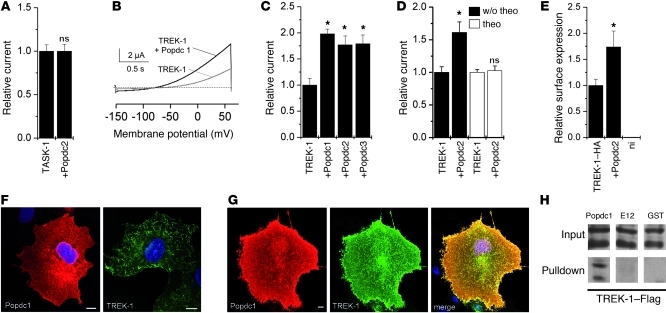
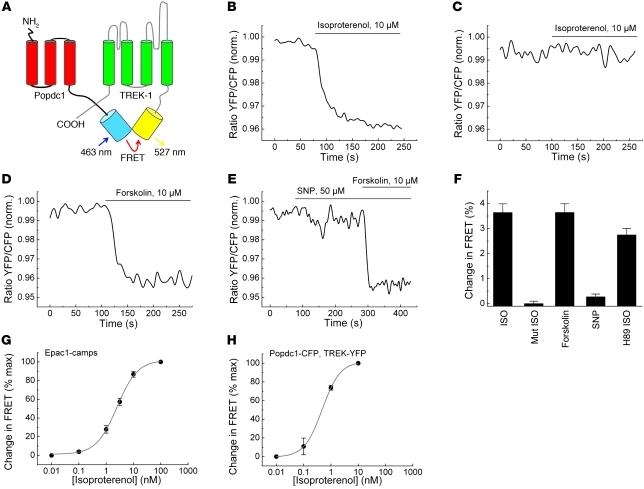
Cardiac pacemaker cells create rhythmic pulses that control heart rate; pacemaker dysfunction is a prevalent disorder in the elderly, but little is known about the underlying molecular causes. Popeye domain containing (Popdc) genes encode membrane proteins with high expression levels in cardiac myocytes and specifically in the cardiac pacemaking and conduction system. Here, we report the phenotypic analysis of mice deficient in Popdc1 or Popdc2. ECG analysis revealed severe sinus node dysfunction when freely roaming mutant animals were subjected to physical or mental stress. In both mutants, bradyarrhythmia developed in an age-dependent manner. Furthermore, we found that the conserved Popeye domain functioned as a high-affinity cAMP-binding site. Popdc proteins interacted with the potassium channel TREK-1, which led to increased cell surface expression and enhanced current density, both of which were negatively modulated by cAMP. These data indicate that Popdc proteins have an important regulatory function in heart rate dynamics that is mediated, at least in part, through cAMP binding. Mice with mutant Popdc1 and Popdc2 alleles are therefore useful models for the dissection of the mechanisms causing pacemaker dysfunction and could aid in the development of strategies for therapeutic intervention.
Figures
Comment in
-
Popeye proteins: muscle for the aging sinus node.J Clin Invest. 2012 Mar;122(3):810-3. doi: 10.1172/JCI62588. Epub 2012 Feb 22. J Clin Invest. 2012. PMID: 22354173 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
