

Loss of the miR-144/451 cluster impairs ischaemic preconditioning-mediated cardioprotection by targeting Rac-1
- PMID: 22354898
- PMCID: PMC3331614
- DOI: 10.1093/cvr/cvs096
Loss of the miR-144/451 cluster impairs ischaemic preconditioning-mediated cardioprotection by targeting Rac-1
Abstract
Aims: While a wealth of data has uncovered distinct microRNA (miR) expression alterations in hypertrophic and ischaemic/reperfused (I/R) hearts, little is known about miR regulation and response to ischaemic preconditioning (IPC).
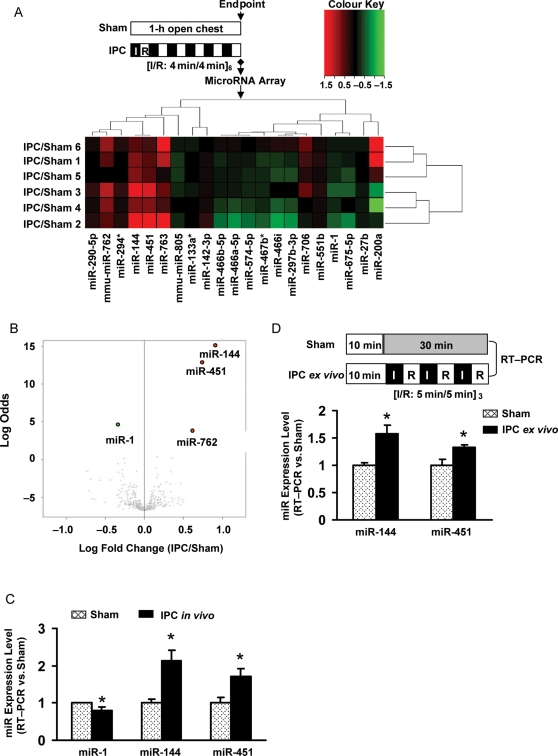
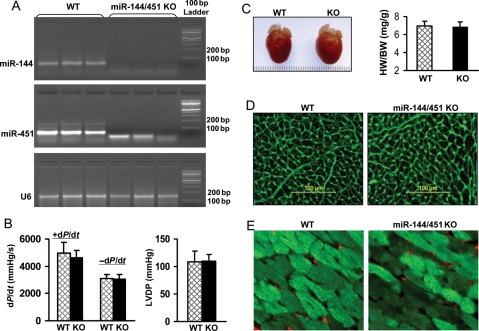
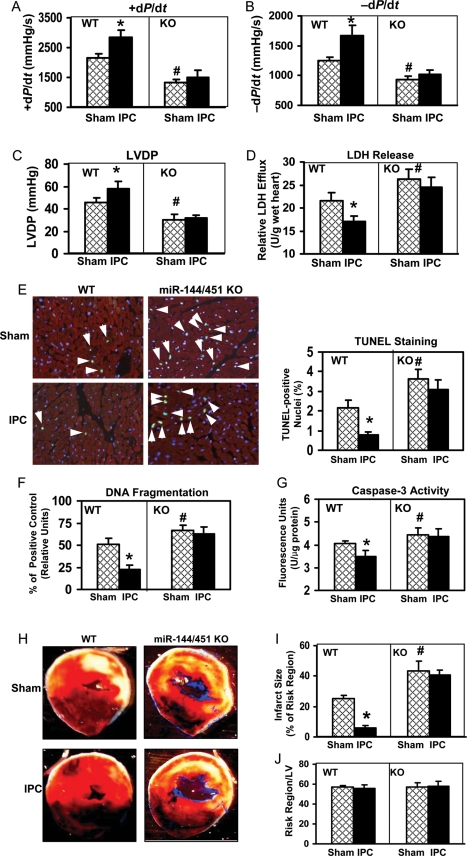
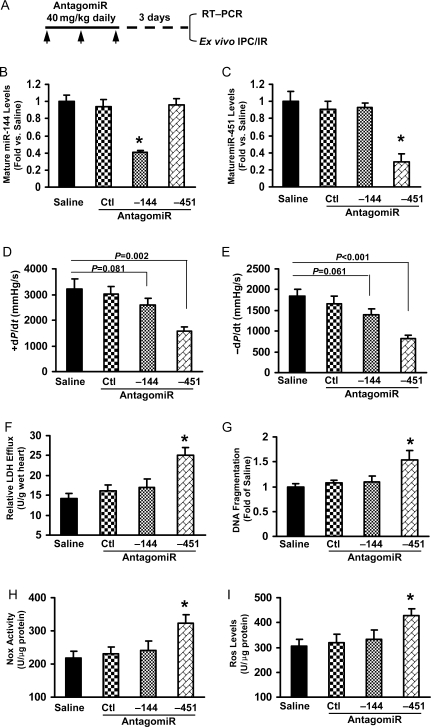
Methods and results: We analysed miRs in murine hearts preconditioned with six cycles of 4 min ischaemia via coronary artery occlusion, followed by 4 min reperfusion in vivo. Both miRs within the miR-144/451 cluster were the most elevated among a cohort of 21 dysregulated miRs in preconditioned hearts, compared with shams. To investigate the significance of this finding, we examined IPC-mediated cardioprotection within a miR-144/451-knockout (KO) mouse model. Wild-type (WT) hearts exposed to IPC followed by I/R (30 min/24 h) showed a smaller infarction size compared with mice treated with I/R alone. In contrast, IPC failed to protect miR-144/451-KO hearts against infarct caused by I/R treatment. Thus, the miR-144/451 cluster is required for IPC-elicited cardioprotection. Rac-1, a key component of NADPH oxidase, was mostly up-regulated in KO hearts among three bona fide targets (Rac-1, 14-3-3ζ, and CUGBP2) for both miR-144 and miR-451. Accordingly, reactive oxygen species (ROS) levels were markedly increased in KO hearts upon IPC, compared with IPC-WT hearts. Pre-treatment of KO hearts with a Rac-1 inhibitor NSC23766 (20 mg/kg, ip) reduced IPC-triggered ROS levels and restored IPC-elicited cardioprotection. Using antagomiRs, we showed that miR-451 was largely responsible for IPC-mediated cardioprotection.
Conclusion: Loss of the miR-144/451 cluster limits IPC cardioprotection by up-regulating Rac-1-mediated oxidative stress signalling.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
