

PK4, a eukaryotic initiation factor 2α(eIF2α) kinase, is essential for the development of the erythrocytic cycle of Plasmodium
- PMID: 22355110
- PMCID: PMC3309761
- DOI: 10.1073/pnas.1121567109
PK4, a eukaryotic initiation factor 2α(eIF2α) kinase, is essential for the development of the erythrocytic cycle of Plasmodium
Abstract
In response to environmental stresses, the mammalian serine threonine kinases PERK, GCN2, HRI, and PKR phosphorylate the regulatory serine 51 of the eukaryotic translation initiation factor 2α (eIF2α) to inhibit global protein synthesis. Plasmodium, the protozoan that causes malaria, expresses three eIF2α kinases: IK1, IK2, and PK4. Like GCN2, IK1 regulates stress response to amino acid starvation. IK2 inhibits development of malaria sporozoites present in the mosquito salivary glands. Here we show that the phosphorylation by PK4 of the regulatory serine 59 of Plasmodium eIF2α is essential for the completion of the parasite's erythrocytic cycle that causes disease in humans. PK4 activity leads to the arrest of global protein synthesis in schizonts, where ontogeny of daughter merozoites takes place, and in gametocytes that infect Anopheles mosquitoes. The implication of these findings is that drugs that reduce PK4 activity should alleviate disease and inhibit malaria transmission.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
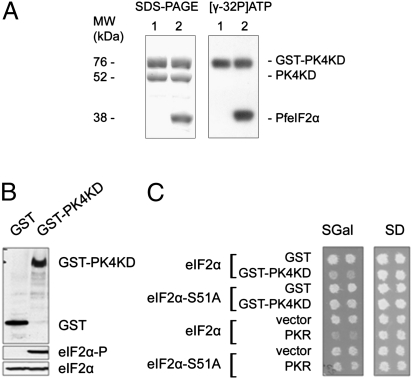
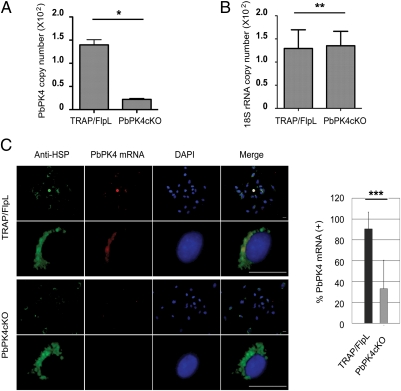
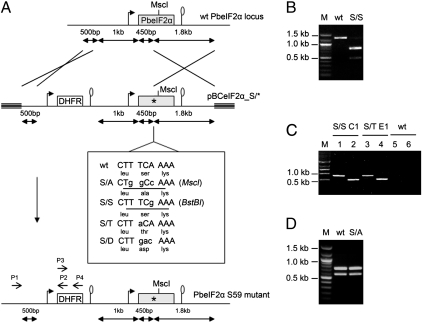
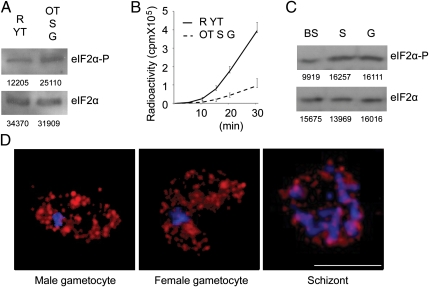
Comment in
-
Parasite physiology: Plasmodium gets the PK4 blood test.Nat Rev Microbiol. 2012 Mar 16;10(4):237. doi: 10.1038/nrmicro2774. Nat Rev Microbiol. 2012. PMID: 22421875 No abstract available.
Similar articles
-
PfeIK1, a eukaryotic initiation factor 2alpha kinase of the human malaria parasite Plasmodium falciparum, regulates stress-response to amino-acid starvation.Malar J. 2009 May 12;8:99. doi: 10.1186/1475-2875-8-99. Malar J. 2009. PMID: 19435497 Free PMC article.
-
The Plasmodium eukaryotic initiation factor-2alpha kinase IK2 controls the latency of sporozoites in the mosquito salivary glands.J Exp Med. 2010 Jul 5;207(7):1465-74. doi: 10.1084/jem.20091975. Epub 2010 Jun 28. J Exp Med. 2010. PMID: 20584882 Free PMC article.
-
The interferon-induced double-stranded RNA-activated protein kinase PKR will phosphorylate serine, threonine, or tyrosine at residue 51 in eukaryotic initiation factor 2alpha.J Biol Chem. 1999 Nov 5;274(45):32198-203. doi: 10.1074/jbc.274.45.32198. J Biol Chem. 1999. PMID: 10542257
-
Small molecule modulators of eukaryotic initiation factor 2α kinases, the key regulators of protein synthesis.Biochimie. 2013 Nov;95(11):1980-90. doi: 10.1016/j.biochi.2013.07.030. Epub 2013 Aug 11. Biochimie. 2013. PMID: 23939221 Review.
-
GCN2 in Viral Defence and the Subversive Tactics Employed by Viruses.J Mol Biol. 2024 Jul 1;436(13):168594. doi: 10.1016/j.jmb.2024.168594. Epub 2024 May 8. J Mol Biol. 2024. PMID: 38724002 Review.
Cited by
-
Translational regulation in blood stages of the malaria parasite Plasmodium spp.: systems-wide studies pave the way.Wiley Interdiscip Rev RNA. 2016 Nov;7(6):772-792. doi: 10.1002/wrna.1365. Epub 2016 May 26. Wiley Interdiscip Rev RNA. 2016. PMID: 27230797 Free PMC article. Review.
-
Artemisinin Action and Resistance in Plasmodium falciparum.Trends Parasitol. 2016 Sep;32(9):682-696. doi: 10.1016/j.pt.2016.05.010. Epub 2016 Jun 9. Trends Parasitol. 2016. PMID: 27289273 Free PMC article. Review.
-
Phosphorylation of eukaryotic initiation factor-2α in response to endoplasmic reticulum and nitrosative stress in the human protozoan parasite, Entamoeba histolytica.Mol Biochem Parasitol. 2019 Dec;234:111223. doi: 10.1016/j.molbiopara.2019.111223. Epub 2019 Sep 27. Mol Biochem Parasitol. 2019. PMID: 31568804 Free PMC article.
-
Plasmodium parasites mount an arrest response to dihydroartemisinin, as revealed by whole transcriptome shotgun sequencing (RNA-seq) and microarray study.BMC Genomics. 2015 Oct 21;16:830. doi: 10.1186/s12864-015-2040-0. BMC Genomics. 2015. PMID: 26490244 Free PMC article.
-
Targeting the ERAD pathway via inhibition of signal peptide peptidase for antiparasitic therapeutic design.Proc Natl Acad Sci U S A. 2012 Dec 26;109(52):21486-91. doi: 10.1073/pnas.1216016110. Epub 2012 Dec 11. Proc Natl Acad Sci U S A. 2012. PMID: 23236186 Free PMC article.
References
-
- Noedl H, et al. Artemisinin resistance in Cambodia: A clinical trial designed to address an emerging problem in Southeast Asia. Clin Infect Dis. 2010;51:e82–e89. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
